

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
"One airway, one disease" is a well-established concept that links the upper and lower airways. Anatomically, these 2 airways have common respiratory epithelium and similar mucosal susceptibility to disease. Epidemiologic studies have consistently shown that asthma and rhinitis often coexist in the same patients. Between 19% and 38% of patients with allergic rhinitis have coexisting asthma, and the prevalence rate of asthma is much higher in patients with allergic rhinitis than in the general population.12 In Korea, the prevalence of bronchial hyper-responsiveness was 55.7% and 25.5% in 6- to 15-year-old patients with allergic and non-allergic rhinitis, respectively.3
Currently, this concept is applied to the middle ear space, given that the middle ear space is an anatomical extension of the airway through the Eustachian tube and capable of mounting an allergic inflammatory response. A previous study showed the incidence of atopy was 24% in the study population comprising patients who had otitis media with effusion (OME) persisting for more than 3 months and unresponsiveness to antibiotics.4 Another study suggested that 35% of patients with recurrent OME have allergic rhinitis.5 Significantly higher numbers of eosinophils and T lymphocytes, as well as significantly higher levels of IL-4 and IL-5 mRNA+ cells, have been reported in patients with atopic OME than in non-atopic controls.6 Furthermore, other studies have shown increased expression of IL-5 and major basic proteins in the middle ear mucosa of patients with OME than that in normal controls, as well as higher levels of eosinophilic cationic proteins in the supernatant of middle ear effusion of atopic patients with OME than in that of non-atopic controls.78 These findings support the concept that the middle ear may be part of the united airway and may behave similarly to the nose or lung under allergic inflammatory conditions. Although allergy has been considered one of the most important factors of OME based on clinical observations, the role of allergy has not been investigated extensively and is easily ignored. In general, the possible integration of the middle ear as part of the united airway concept will have major clinical implications for the diagnosis and management of allergic airway disease.
Dermatophagoides pteronyssinus (Der p) and Dermatophagoides farinae (Der f) are the most common house dust mite (HDM) species, which produce allergens that are widely distributed in Korea. Recently, Der p and Der f have been shown to activate toll-like receptors (TLRs) 2 or 4 and thereby stimulate the innate immune response. TLRs recognize repetitive patterns in diverse microbes, including gram-positive and gram-negative bacteria and viruses, and they are key components of innate immunity.910 In the innate immune response, airway epithelial cells aid the immune system by inducing recruitment of immune-competent cells to local tissues and thereby modulating their activity.11 In addition, lipopolysaccharide (LPS), a major component of the outer membrane of gram-negative bacteria, is a virulence factor that can induce inflammatory responses. LPS may induce the release of multiple pro-inflammatory cytokines and chemokines from airway epithelial cells, and this initial inflammatory response plays a key role in the containment of infection. The inflammatory response induced by LPS in epithelial cells has been linked to TLR 4.12
The airway mucosa is lined with ciliated columnar epithelium interspersed with goblet cells. Accumulation of secretions and mucostasis in the airway mucosa due to allergic airway disease can alter the viscoelasticity of the mucus, ciliary motility, and epithelial defense mechanisms, thereby increasing the susceptibility to various stimuli, such as bacteria and viruses.13 Viruses are clinically linked to allergy: viral infection can induce asthma/allergic exacerbations, and viral clearance and persistence of symptoms are prolonged in allergic individuals.141516 Conversely, chronic allergic inflammation may impair anti-viral reaction to acute rhinovirus infection, as indicated by the suppression of IFN-α, IFN-γ, and IL-12 induction.17 Meanwhile, airborne LPS might adversely affect asthmatic individuals by enhancing an established airway inflammation and airway obstruction.1819
Considering the above findings, it is natural to think that epithelial cells of the middle ear might be involved in recognizing and formulating an innate immune response to Der f and LPS, and a kind of reciprocal effect may be developed by subsequent exposure to Der f and LPS. Although allergy or allergen sensitization could be a concealed, potent stimulant of inflammation in sinus and middle ear disease, the effect of allergen sensitization on infection and its mechanism in airway epithelium is not clearly understood. Moreover, although allergy has been implicated in OME, the role of middle ear epithelial cells in the innate immune response has not been characterized. Thus, this study was design to investigate whether allergen pre-sensitization within airway mucosa can affect subsequent viral or bacterial infection. For this, we performed in vitro experiments using primary human middle ear epithelial cells (HMEECs) and assessed the signaling pathways activated by Der f and LPS as well as the production of pro-inflammatory cytokines and the expression of the mucin (MUC) gene.
MATERIALS AND METHODS
Reagents
LPS, SB203580, and Bay were obtained from Sigma-Aldrich (Munich, Germany). Der f crude body extract was purchased from Arthropods of Medical Importance Resource Bank (Seoul, Korea). Other chemicals used were of the purest grade available from Sigma (St. Louis, MO, USA).
Cell culture and stimulation
HMEECs (kindly provided by Dr. David J. Lim, House Ear Institutes, Los Angeles, CA, USA) were maintained in a mixture of Dulbecco's modified Eagle's medium (DMEM, Invitrogen, Carlsbad, CA, USA) and bronchial epithelial basal medium (Lonza, Walkersville, MD, USA) (1:1).202122 Cells were grown to 60% confluence in 100-mm culture plates and kept at 37℃ in a carbon dioxide-enriched (95% air, 5% CO2) humidified atmosphere.
HMEECs were seeded onto 60-mm culture plates, with 2.2×105 cells per well for the experimental condition. Predeter-mined non-cytotoxic doses of Der f or LPS by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay (data not shown) were applied to HMEECs. By performing the pilot study using real-time PCR, treatment with 1.0 µg/mL LPS for 24 hours was identified to be the optimal and effective in that it not only elicits a moderate level of inflammatory response but also has an augmentative effect following the administration of 2.0 µg/mL Der f. Cells were then stimulated in several ways: 1) Der f 2.0 µg/mL only for 48 hours (Df48h); 2) LPS 1.0 µg/mL only for 24 hours (LPS24h); 3) Der f 2.0 µg/mL for 48 hours and add LPS 1.0 µg/mL for 24 hours (Df48h/LPS24h); 4) LPS 1.0 µg/mL for 48 hours and add Der f 2.0 µg/mL for 24 hours (LPS48h/Df24h); or 5) Der f 2.0 µg/mL and LPS 1.0 µg/mL both simultaneously for 48 hours (Df48h/LPS48h) (Fig. 1).
Real-time polymerase chain reaction (qRT-PCR)
Total RNA was isolated from the epithelial cells using TRIzol (Invitrogen) as per the manufacturer's instructions and reverse transcribed into cDNA using the Quantitect Reverse Transcription kit (Qiagen, Venlo, Netherlands). Then qRT-PCR analyses were performed using a 7,500 FAST qRT-PCR System (Applied Biosystems, Foster City, CA, USA). Each reaction mixture contained 10 µL of SYBR® Green PCR Master Mix (Applied Biosystems), 4 pmol each of the forward and reverse primers, and 1 µL of cDNA in a final volume of 20 µL. Reaction mixtures were incubated at 95℃ for 5 minutes to activate FastStartTaq DNA Polymerase, followed by 40 cycles of amplification. Data were analyzed using Sequence Detection Software version 1.9.1 (Applied Biosystems). Target mRNA expression was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression and calculated using the comparative Ct method. Primers used in this study are shown in Table.
RNA interference and plasmid DNA transfection
Cells were transfected with control small interfering (si) RNA, TLR 4 siRNA or myeloid differentiation (MD)-2 siRNA, and 10 µL of Lipofectamine RNAiMAX (Invitrogen) in 60-mm plates as per the manufacturer's protocol. The plasmid pFlag-CMV1-hMD2 was a kind gift from Doug Golenbock (Addgene plasmid #13028). After cells were washed with OPTI-MEM (Gibco, Carlsbad, CA, USA), DNA was transfected to cells using X-treme GENE HP-DNA transfection reagent (Roche Diagnostics, Indianapolis, IN, USA) as per the supplier's protocol. After 4 hours of incubation, the medium was exchanged for a complete medium containing 10% serum and antibiotics. Cells were incubated for an additional 24 hours and treated with Df or LPS. Cell viability was measured by light microscopy, and the gene-silencing or expression efficacy was evaluated by evaluating the mRNA levels. The siRNAs were NM_138554.2 for TLR4 and NM_014364.2 for MD-2 (Bioneer, Daejeon, Korea).
Western blot analysis
At specific time points after the Der f or LPS treatment, the medium was removed, and cells were washed with phosphate-buffered saline (PBS; 10 mM, pH 7.4). Cells were then lysed with lysis buffer (50 mM Tris pH 7.7, 150 mM NaCl, 1% NP-40, 5 mM EGTA, 50 mM -glycerophosphate, 20 mM NaF, 1 mM Na3VO4, 2 mM phenylmethylsulfonyl fluoride, 10 mg/mL leupeptin, and 10 mg/mL aprotinin) and incubated for 20 minutes at 4℃. Briefly after sonication, cells were centrifuged at 13,000×g for 10 minutes at 4℃. The supernatant containing the total cell lysate was collected. Protein concentration of the lysates was measured using the Bio-Rad Protein Assay (Bio-Rad, Hercules, CA, USA). Equal amounts of protein were mixed with sample buffer (62.5 mM Tris–HCl [pH 6.8], 1% sodium dodecyl sulfate, 2.5% glycerol, 0.5% 2-β-mercaptoethanol, and bromophenol blue), boiled for 5 minutes, and separated by electrophoresis on 10%-12% Tris-HCl gels. Protein content of the gels was transferred to a PVDF membrane (Amersham, Buckinghamshire, UK), and the membranes were blocked with TBS-T (20 mM Tris, 500 nM NaCl, with 0.1% Tween-20) containing 5% (w/v) skim milk for 1 hour at room temperature. Membranes were probed with antibodies against MUC 8 (Sigma–Aldrich, St. Louis, MO, USA), TLR-4, p-p38, p38, p-CREB (Santa Cruz Biotechnology, Dallas, TX, USA) and GAPDH (Santa Cruz Biotechnology) followed by peroxidase-conjugated anti-mouse IgG or anti-rabbit IgG (Jackson Immuno Research, West Grove, PA, USA).Membranes were developed using the enhanced chemiluminescent analysis system (SuperSignal® West Pico Chemiluminescent Substrate, Pierce, Waltham, MA, USA) and the signal was captured using an image reader (LAS4000; Fuji Photo Film, Tokyo, Japan). Results were obtained from three independent experiments.
Immunofluorescence
HMEECs were transfected with siRNA-TLR4, MD-2, and control, and treated with Der f 24h and then with LPS24h on a cover slide in 12-well plates. Cells were rinsed 3 times with PBS, fixed with 4% paraformaldehyde for 10 minutes at room temperature, and rinsed again. Cells were then blocked with 1% bovine serum albumin, followed by the addition of the primary antibodies anti-Der f and anti-TLR4. After extensive washing with PBS, fluorescein isothiocyanate-conjugated IgG was added. Following incubation, the slides were rinsed, mounted, and viewed at 488 nm on a confocal microscope (FV1000, Olympus, Japan).
Statistical analysis
Statistical analyses were performed using SPSS for Windows (Ver. 12.0, SPSS Inc., Chicago, IL, USA). All data are expressed as mean ± standard deviation (SD). A one-way analysis of variance (ANOVA) was used to determine statistical significance differences between control and groups at each time or dose point. Scheffe's F test was used to correct for multiple comparisons when statistically significant differences were identified in the ANOVA. A P value of <0.05 was considered statistically significant.
RESULTS
Induction of MUC and pro-inflammatory cytokine gene mRNAs expression in HMEECs stimulated by Der f and LPS
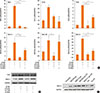
Treatment with Df48h increased the level of MUC 8 mRNA expression. The expression level was significantly augmented when cells were exposed to Df48h/LPS24h. We further analyzed the interaction of Der f and LPS under different treatment conditions. When we treated the cells in either with 1) LPS48h/Df24h or 2) Df48h/LPS48h, MUC8 mRNA expression did not reach the levels induced by Df48h/LPS24h (Fig. 2A). With regard to the MUC 4, 5AC and 5B mRNA expression levels increased in a similar fashion, showing the greatest increase in the cells treated Df48h/LPS24h without statistical significance (Fig. 2B). This pattern was confirmed at the protein level. Immunoblot assay showed that Df48h/LPS24h synergistically elevated the MUC 8 protein production; however, this was not observed in Df48h or LPS24h (Fig. 3A). Next, we analyzed the magnitude of the synergistic effect of both Der f and LPS by evaluating the MUC 8 and TLR 4 protein expression levels, because both Der f and LPS are known to bind to TLR 4 to trigger the inflammatory signaling pathway. To further demonstrate the synergistic effect of Der f and LPS, we treated HMEECs with LPS 2, 4, 6, 8, and 10 µg/mL each for 24 hours, and then compared the intensity of MUC8 and TLR 4 proteins produced by Df48LPS24 treatment. Df48h/LPS24h showed a higher intensity than that produced by LPS24h only (>10 µg/mL) (Fig. 3B). In terms of mRNA expression of pro-inflammatory cytokines, such as IL-1ß, GM-CSF, IL-33, and TNF-α, the augmented pattern was also similar to that of MUC, although they did not reach statistical significance (Fig. 4). These results suggest a possibility that Der f and LPS can act synergistically.
Role of TLR4 in the induction of the MUC gene in Der f- and LPS-stimulated HMEECs
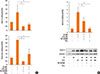
To elucidate the mechanism underlying the difference in mucin gene expression in cells treated with LPS48h/Df24h or Df24h/LPS48h, we further determined the roles of TLR 4 and MD-2. We therefore transfected HMEECs with siRNAs to knock down the expression of TLR4 or MD-2. Suppression of TLR4 and MD-2 by each siRNA were confirmed by real-time PCR. CD14 expression was also suppressed by both siRNA without statistical significance (Fig. 5A, upper panel). Expressions of MUC 4, 5B and 8 mRNAs by the treatment with Df48h/LPS24h decreased significantly upon siRNA-TLR4 transfection. However, a significant amount of mRNAs were still expressed in siRNA-MD-2 transfected HMEECs (Fig. 5A, lower panel). Immunoblot analysis further confirmed decreased or sustained increase in MUC 8 protein signals in siRNA-TLR4- and siRNA-MD-2-transfected cells, respectively (Fig. 5B). These results indicated that TLR 4 could be partially activated in the absence of MD-2 protein.
Alternatives to MD-2 for MUC gene induction in Der f- and LPS-stimulated HMEECs
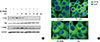
Our above mentioned results suggest the possibility of Der f being a functional substitute for the MD-2 protein; thus, the TLR 4 signal could be activated without the help of MD-2 protein. Therefore, to determine the exact role of MD-2 in Der f and LPS synergism, a mammalian expression vector containing MD-2 (pCMV1-hMD-2) was transfected into HMEECs for transient expression studies. MD-2 was over-expressed in HMEECs subsequently exposed to LPS for 24 hours, which showed a higher MUC 8 protein signal density than those exposed to LPS only and similar MUC 8 protein signals compared to those exposed to Df48h/LPS24h (Fig. 5C). Moreover, we conducted the co-immunofluorescence staining of TLR 4 and Der f in siRNA-treated cells. Der f and TLR 4 were co-localized not only in the siRNA-negative control transfected cells but also in the siRNA-MD-2 transfected cells (Fig. 6A and B). These results suggest that the MD-2 protein may play an important role in the pro-inflammatory Der f and LPS synergism.
p38 MAPK and CREB phosphorylation inhibition and NF-κB translocation in Der f-and LPS-stimulated HMEECs
To further investigate the mechanism underlying the synergistic functions of Der f and LPS, we treated cells with various inhibitors, such as SB203580 (a bicyclic inhibitor of p38), PD98059 (a specific inhibitor of MAPK/MEK-1), Bay (a NF-κB inhibitor), LY294002 (PI3K inhibitor), and a JNK inhibitor (data not shown). Cells were treated with each inhibitor 2 hours prior to LPS treatment. Of these, the mRNA expression levels of MUC 2, 5AC, and 8 decreased significantly under SB203580 or Bay treatment (Fig. 7A). MUC 8 protein signal intensity was lower in SB203580-or Bay-treated cells than in Df48h/LPS24h cells and most lowest in Df48h/LPS48h cells (Fig. 7B). This suggests that p38 MAPK and NF-κB transcriptional factors may be engaged in the synergism of Der f and LPS. The p-p38 signal post LPS treatment peaked at 5 minutes, and the expression of p-p38 and p-CREB was inhibited by SB203580 (Fig. 8A). GAPDH was constitutively expressed and not affected by SB203580. Moreover, NF-κB translocation was seen in Df48h/LPS24h by confocal microscopy (Fig. 8B).
DISCUSSION
We investigated the synergistic effect of Der f and LPS on MUC gene expression and pro-inflammatory cytokine production in HMEECs. LPS is known to induce and potentiate MUC gene expression;3 however, the interaction between LPS and Der f has not been studied so far. Before starting the experiment, to rule out possible endotoxin contamination of Der f crude body extract, we first measured the endotoxin level via the LAL method (Toxin Sensor chromogenic LAL Endotoxin assay kit, Gen Script, Piscataway, NJ, USA). The endotoxin level in Der f crude body extract was found to be 0.16 EU/mL, which was considered negligible. In this study, Der f and LPS synergistically induced MUC 8 gene expression and pro-inflammatory cytokines, and this effect depends on the sequence of exposure, i.e., it only occurs when Der f stimuli precedes LPS stimuli. Additionally, pre-treated Der f is likely to function as an MD-2 protein and induce p-p38 and p-CREB expressions. These findings provide evidence for the interaction between 2 environmental stimuli associated with allergic airway disease and infectious diseases in the airway epithelium.
We measured mucin gene production as the final product because it is one of the major molecules that the airway epithelium produces in large amounts in response to several stimuli, such as TNF-α, IL-1β, LPS, oxidative stress, and neutrophil elastase, and because it has been implicated in numerous airway diseases.24 Mucins are broadly classified as either cell membrane-bound or secreted; notably, MUC2, MUC5AC, MUC5B, MUC6, MUC7, MUC8, MUC9, and MUC19 are categorized as secreted mucins.2526 MUC5AC and MUC5B are predominant in the sinus and nasal cavity, respectively, and considered primarily responsible for the gel-like characteristics of mucoid middle ear fluids.27282930 The MUC 8 gene is over-expressed in chronic rhinosinusitis and nasal polyp epithelium, and its expression levels are also increased by inflammatory mediators in nasal epithelial cells.3132 Airway mucus obstruction is a shared feature among lower airway diseases, such as cystic fibrosis, asthma, and chronic obstructive lung disease.33 Hence, investigating the mucin gene expression in HMEECs is considered an appropriate approach to evaluating the "extended one airway concept."
Interactions between allergens, viruses, bacteria and/or other pro-inflammatory stimuli have been previously reported.3 The synergistic effects of human rhinovirus and nitric oxide 2 on IL-8 release and an additive effect on ICAM-1 expression have been demonstrated.34 The interaction between the human rhinovirus or respiratory syncytial virus and Der p I has been studied, and it has been found that they synergistically induce IL-8 expression in bronchial epithelial cells.35 With regard to the interaction between bacteria and allergens, a previous study has shown that systemic LPS administration suppresses early and late allergic reactions in vivo via the TLR4-dependent pathway that triggers nitric oxide synthase 2 activity in a murine model of asthma;36 the generation of Th1 or Th2 responses is found to be dependent on the dose of LPS. At a low LPS level, inhaled LPS induces a Th2 response to inhaled antigens, whereas at a high level, inhaled LPS results in a Th1 response to the antigen.37 In an HDM allergen-evoked asthma murine model, LPS dose-dependently inhibits HDM-induced eosinophilic recruitment into the lungs, mucus production in the airways, and production of Th2 cytokines (IL-4, IL-5, IL-10, and IL-13).38 However, the interaction between Der f and LPS at a cellular level has not been addressed previously, especially in the middle ear epithelium in vivo or in vitro, despite being frequently implicated in middle ear disease pathogenesis.
It is well known that co-molecules, such as MD-2, CD14, myD88, and LPS-binding protein are needed to activate the TLR 4 signaling pathway; in particular, MD-2, a glycoprotein co-expressed with TLR4 at the surface of various cell types, has received recent attention. Theoretically, to transmit the LPS-driven inflammatory signals, LPS bound to LBP is shuttled to CD14, which transfers the LPS to an MD-2 molecule. Activation of MD-2 occurs in 2 ways: it combines with endogenously expressed TLR-4 to increase the surface expression of TLR-4 and MD-2,39 and participates directly in endotoxin recognition by extracting the endotoxin from monomeric endotoxin:CD14 complex.40
Previous studies demonstrate that MD-2 is essential for LPS-driven inflammatory responses, and that lack of MD-2 protein expression in human corneal epithelial cells account for the lack of response to LPS exposure in the cornea.41 MD-2 binds to TLR 4 and greatly enhances the response of TLR4-transfected cells to LPS exposure.42 Patients with the single nucleotide polymorphisms of MD-2 promoter increase mRNA expressions of MD-2 and tended to have high levels of IL-1β, IL-6, IL-8, IL-10, and tumor necrosis factor (TNF)-α after Der p 2 and LPS stimulation.43 These facts prompted us to investigate whether the MD-2 protein might have a specific role in allergen and LPS synergism in HMEECs. In our study, to evaluate the role of MD-2, we knocked-down the MD-2 gene by transfecting siRNA-MD-2. Interestingly, the production of MUC genes was not completely decreased in the absence of MD-2. Moreover, pre-treated Der f did not significantly alter the MD-2 mRNA expression level (data not shown). Overexpression of the MD-2 gene using the pCMV-hMD-2 vector had a similar induction effect of MUC 8 protein when followed by LPS exposure for 24 hours. This implies that Der f may combine with LPS, TLR4, and CD14, and have a role in endotoxin recognition in cases of deficiency of the MD-2 protein. Notably, the crystal structures of MD-2 and Der p 2 exhibit structural homology with 2 anti-parallel β-pleated sheets stabilized by disulfide bonds.444546 In an in vivo study, airway sensitization and Der p 2 challenge resulted in allergic asthma in wild-type and MD-2-deficient mice, but not in TLR4-deficient mice.47 A previous study stated that human airway epithelium internalizes Der p2, which in turn, increases the endogenous cellular expression of MD-2.48 Der p 2 has not only structural homology but also functional homology with MD-2, facilitating signaling through direct interactions with the TLR4 complex and reconstituting LPS-driven TLR4 signaling in the absence of MD-2.47 Taken together, these facts suggest that Der f may have functional homology with MD-2.
Meanwhile, there are several additional factors that may influence augmented mucin gene expression induced by Der f and LPS. Notably, at least 3 different microbial pathogen-associated molecular patterns can be detected in mite feces and/or in the mite environment: LPS, β-glucan, and chitin. Both LPS and β-glucan mediate innate immunity through reactive oxygen species production and can subsequently affect the mucin gene production.49 The endotoxin level of Der f crude body extract used in this study was 0.16 EU/mL, and thus the contribution might not be significant. Chitin can polarize Th1, Th2, and Th17 immune responses, and recruit IL-4 positive innate immune cells, including eosinophils and basophils;5051 however, its precise contribution in the HDM-LPS synergism remains to be elucidated.
HDM allergenic proteins can be categorized into 4 main families: proteases, proteins displaying affinity for lipids, non-proteolytic enzymes, and non-enzymatic components.52 In the context of innate immunity activation, all these protein and non-protein compounds could putatively participate in stimulation. Among them, group 2 HDM allergen is described as an MD2-like lipid binding protein based on its structural/sequence homology,53 supporting the hypothesis of our study.
Finally, the lack of synergism when the HMEECs were treated with Der f and LPS simultaneously (Df48h/LPS48h) or LPS48h/Df24h may be attributed to the following aspects. First, it has been noted that Der f binds to LPS with a molar ratio of 1:1.54 Thus, when treated simultaneously, a fraction of these molecules may bind before reaching and binding to TLR4 and thus cannot activate the signaling pathway as expected. Second, an insufficient number of TLR 4 on the cell surface or endogenous MD-2 protein may interact with Der f and LPS when they are added at the same time. Third, the "hygiene hypothesis" states that the high levels of exposure to bacterial products, such as LPS, early in life are inversely correlated with the development of atopy and allergic disease.555657 Thus, it is thought that LPS exposure leads to the exhaustion of TLR 4 and the development of a counter-regulatory response.58 Finally, unlike in LPS48h/Df24-treated cells, Der f would act like MD-2 protein in Df48h/LPS24h-treated cells, more signal activity could be triggered.
In summary, these experiments demonstrate that Der f and LPS can act synergistically when Der f exposure precedes LPS exposure in HMEECs. Based on our findings, the likely underlying mechanism is that Der f has functions similar to those of MD-2, combining with TLR-4 and participating in endotoxin recognition. Such synergy suggests its important role in the development of OME in patients with concealed allergen-sensitized airway epithelium. Thus, targeted inactivation of innate immune signals to allergen exposure may be useful for the development of specific therapeutics for otitis media in concealed allergic sensitization.






 XML Download
XML Download