

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Pigment epithelium-derived factor (PEDF) is a 50 kDa secreted glycoprotein with multiple functions, including anti-inflammation, antiangiogenic and antifibrosis properties.123 PEDF has been found to inhibit NF-ΚB activation and the expression of several proinflammatory genes such as intercellular cell adhesion molecule-1 (ICAM-1), tumor necrosis factor alpha (TNF-α) and matrix metalloproteinases.456 PEDF has also been shown to participate in the pathogenesis of allergic rhinitis.7 These functions suggest that PEDF may be a potential target in the prevention and treatment of allergic asthma.
Bronchial asthma is a chronic airway inflammatory disease associated with airway hyperresponsiveness (AHR) and airway remodeling. Airway remodeling refers to structural changes, including mucous gland hyperplasia, smooth muscle proliferation, subepithelial fibrosis, and vascular changes.89 Vascular endothelial growth factor (VEGF) is believed to be involved in the pathogenesis of allergic asthma, particularly airway remodeling.10 VEGF expression is regulated by several stimuli, including TNF-α, interleukin (IL)-1β, and lipopolysaccharides (LPS).11 Recent studies have shown that PEDF can negatively regulate VEGF expression.1213 We hypothesize that PEDF treatment may down-regulate VEGF expression and may lead to the inhibition of airway inflammation and airway remodeling.
The purpose of this study was to define the effects of PEDF on allergic airway inflammation and airway remodeling in BALB/c mice and in BEAS-2B human bronchial epithelial cells with in vivo and in vitro experiments, respectively. Our results clearly show that PEDF inhibits allergic airway inflammation and airway remodeling, at least in part, by suppressing VEGF expression.
MATERIALS AND METHODS
Animals
Six- to eight-week-old female BALB/c mice (each weighing approximately 20 g) were purchased from Shanghai Laboratory Animal Inc. (Shanghai, China). All experimental animals were utilized under protocols approved by the Institutional Animal Care and Use Committee of Nanjing Medical University and the institutional animal ethics committee (Nanjing, China).
Antigen sensitization, challenge, and treatment
Thirty two mice were randomly divided into the control, OVA, PEDF low and PEDF high groups. On Days 0 and 14, the mice in the OVA, PEDF low, and PEDF high groups were immunized by intraperitoneal injection of 100 µg of chicken egg ovalbumin (OVA, Grade V; Sigma, St Louis, MO, USA) emulsified in 100 µL of aluminum hydroxide gel (InvivoGen, San Diego, CA, USA). On day 21, the mice were placed in a Plexiglas box (29×22×18 cm) and were airway challenged with 1% aerosolized OVA for 30 minutes per day, 3 days per week, for a period of 8 weeks. The mice in the PEDF low and high groups were given injections via the tail vein with 50 or 100 µg/kg body weight of recombinant PEDF protein13 (Peprotech, Rocky Hill, NJ, USA) before each OVA challenge. The mice in the control group received sensitization and airway challenge with phosphate-buffered saline (PBS) instead of OVA.
Airway hyperreactivity measurement
Airway responsiveness to acetylcholine chloride (ACh) was measured 24 hours after the last OVA challenge with an AniRes 2005 animal lung function analysis system (SYNOL High-Tech, Beijing, China) as previously described.16 Mice were anesthetized with an intraperitoneal injection of pentobarbital sodium (70 mg/kg). The trachea was then surgically exposed, and a plastic tube with an internal diameter of 4 mm was inserted into the trachea connected to a computer-controlled ventilator. A 27-gauge needle was inserted into the tail vein for ACh administration. The respiratory rate and the tidal volume were preset at 90 breaths/min and 6 mL/kg, respectively. Progressively increasing doses of ACh (10, 30, 90, and 270 µg/kg) were administered intravenously with a microinfusion pump (36 mL/min) via the caudalis vein. Data were obtained, and the maximum values of lung resistance (RL) were used to express changes in airway hyperreactivity.91011
Analysis of bronchoalveolar lavage fluid and serum
After measurement of airway hyperreactivity, the retro-orbital puncture method was used to collect blood samples. Serum samples were collected after centrifuging at 1,000 g at 4℃ for 15 minutes, and plasma was stored at -70℃ until analysis. Airway lumina were washed 3 times with 0.5 mL volumes of saline. Bronchoalveolar lavage fluid (BALF) was centrifuged at 1,000 g at 4℃ for 15 minutes, and upper fluid samples were collected for detection by enzyme-linked immunosorbent assay (ELISA). The cell pellets were suspended for total inflammatory cell counts with a hemocytometer. The smears of BALF cells were stained with Wright's stain for differential cell counts. The cells in the BALF were counted by 2 independent investigators in a single-blind study analyzing at least 200 cells each from 4 different random locations using a microscope. Commercially available ELISA kits were used to evaluate the levels of the Th2 cytokines IL-4 (Jingmei Biotech, Shanghai, China), IL-5, IL-13, transforming growth factor-beta1(TGF-β1), and VEGF (R&D Systems, Minneapolis, MN, USA) in BALF and OVA-specific serum IgE (Shibayagi, Gunma, Japan).
Lung histology and immunohistochemistry
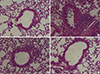
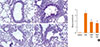
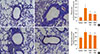
The left lung was fixed in 4% paraformaldehyde overnight at 4℃ and then embedded in paraffin. Paraffin sections (5 µm) were stained with hematoxylin and eosin (H&E) to detect eosinophilic infiltration, periodic acid-Schiff (PAS) to assess mucus-secreting goblet cells, and Masson's trichrome stain to evaluate the peribronchiolar collagen layer or processed for α-smooth muscle actin (SMA) immunohistochemistry to estimate airway smooth muscle hypertrophy. Histological analyses were performed by pathologists blinded to the treatment groups. Scoring for mucus production was quantified as previously described15: 0, no goblet cells; 1, <25% of the epithelium; 2, 25%-50% of the epithelium; 3, 50%-75% of the epithelium; and 4, >75% of the epithelium. The area of peribronchial trichrome staining in the mouse lung is expressed as the area of trichrome staining per µm length of the basement membrane (BM) of bronchioles. At least 10 bronchioles with internal diameters of 150-200 µm in each slide were counted.
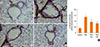
Lung sections were treated in xylene and rehydrated in graduated alcohol solutions. Endogenous peroxidase activity was blocked with 3% H2O2. Specimens were flooded with 5% normal goat serum to prevent the nonspecific absorption of immunoglobulin; then, specimens were incubated with anti-α-SMA monoclonal antibody (DAKO, Glostrup, Denmark) at a dilution of 1:50 for 90 minutes. A substitution of each primary antibody with PBS was used as a negative control. The slides were incubated overnight at 4℃, rinsed 3 times with PBS, and then incubated with peroxidase-labeled secondary antibody (DAKO, Glostrup, Denmark) for 30 minutes at room temperature. The sections were washed again with PBS, followed by diaminobenzidine staining. The sections were then counterstained with hematoxylin, dehydrated, and observed. Data are expressed as the area of α-SMA immunostaining per µm length of the BM of bronchioles with internal diameters of 150-200 µm.
Measurement of hydroxyproline content
A hydroxyproline assay was used to determine the total collagen content of the right lungs using a commercially available hydroxyproline detection kit (Nanjing Jiancheng Biotechnology, Nanjing, China). Briefly, frozen right lung tissues were homogenized in saline containing 0.1 mol/L phenylmethylsulfonyl fluoride. Then, the homogenized lung tissues were hydrolyzed in 6 mol/L HCl. The hydroxyproline concentration was measured as previously described.17
Cell culture and treatment
BEAS-2B human bronchial epithelial cells were cultured in 6-well round-bottom plates supplemented with 4 mM L-glutamine, 20 µg/mL streptomycin, 100 U/mL penicillin, and 10% fetal bovine serum (all from HyClone, Logan, UT, USA). After starvation, the cells were cultured with 500 ng/mL IL-1β (R&D Systems, Minneapolis, MN, USA) in the presence or absence of different doses of recombinant PEDF protein (50,100 ng/mL, Peprotech, USA).1819 The supernatants were collected after 48 hours, and VEGF levels were assayed using ELISA kits.
Western blot analysis of VEGF in the lung tissue
The right lung tissues from each group were washed with ice-cold PBS and homogenized, and the lysates were prepared in RIPA lysis buffer and centrifuged at 12,000 g at 4℃ for 15 minutes. Equal amounts (30 µg) of proteins were loaded onto 8% SDS-PAGE gels. Subsequently, the proteins were electrotransferred (100 V for 2 hours). The membrane was blocked for 1 hour with Tris-buffered saline containing 0.05% Tween 20 (TBST) plus 5% skim milk and then incubated with 1:1,000 anti-VEGF (Abcam, Cambridge, UK) or 1:1,000 anti-GAPDH (Cell Signaling Technology Inc., Beverly, MA, USA) overnight at 4℃. The membrane was washed 3 times with TBST and then incubated with HRP-conjugated goat antirabbit IgG for 1 hour at room temperature. The membrane was washed 3 times with TBST again and detected with an ECL detection system (Pierce Biotechnology, Rockford, IL, USA).
RESULTS
PEDF decreases AHR in chronic experimental asthma
To determine the effects of PEDF on airway reactivity, lung resistance (LR) was evaluated in anesthetized mice by invasive whole-body plethysmography. There were no significant differences in baseline airway resistance between the 4 groups. The LR generated by the administration of Ach at doses from 30 to 270 µg/kg was dramatically increased in the OVA group compared to the control group. RL was significantly decreased in the recombinant PEDF treated group (50 or 100 µg/kg bodyweight) compared to the OVA group, implying that in vivo inflammation-mediated airway pathology was alleviated. Recombinant PEDF reduced AHR in a dose-dependent manner (Fig. 1A).
PEDF attenuates chronic airway inflammation
To determine the effects of PEDF on airway inflammation, the amount and classification of inflammatory cells in the BALF were evaluated. Few inflammatory cells were detected in saline-sensitized and saline-challenged control mice. The mice in the OVA group displayed a marked increase in the number of total inflammatory cells and eosinophils in the BALF. Treatment with PEDF significantly decreased the numbers of eosinophils and total inflammatory cells in BALF in a dose-dependent manner (P<0.05, Fig. 1B). In addition, HE staining showed a marked reduction in the number of total inflammatory cells around the airway and the blood vessels in PEDF-treated mice compared to the OVA group (Fig. 2).
PEDF reduces Th2 cytokines and TGF-β1 levels in BALF
To determine the effects of PEDF on Th2 cytokines and TGF-β1, we measured the levels of T-helper type 2 (Th2) cytokines, such as IL-4, IL-5, IL-13, and TGF-β1 in BALF. IL-4, IL-5, IL-13, and TGF-β1 in BALF were dramatically increased in the OVA group compared to the control group (P<0.05, Fig. 3). Administration of PEDF significantly decreased Th2 cytokine and TGF-β1 levels in BALF in a dose dependent manner compared to the OVA group (P<0.05, Fig. 3).
PEDF decreases OVA-specific IgE in serum
To evaluate the effect of PEDF on IgE in serum, we also measured the level of OVA-specific IgE in serum. OVA-specific IgE in serum was obviously enhanced in the OVA group compared to the control group (P<0.05, Fig. 3), treatment with PEDF significantly inhibited the level of OVA-specific IgE in a dose dependent manner (P<0.05, Fig. 3).
PEDF inhibits airway remodeling in a chronic asthma model
To determine the effects of PEDF on airway remodeling changes, we determined the effects of PEDF on goblet cell metaplasia, smooth muscle proliferation, and subepithelial fibrosis. PAS staining was used to identify mucus-containing cells in the airway epithelium (Fig. 4A-D). Significant goblet cell hyperplasia and mucus hypersecretion were found in the OVA mice compared to the control group (P<0.05, Fig. 4E). This increase was evidently inhibited in mice treated with PEDF. Furthermore, PEDF reduced the number of goblet cells in the lumen of the bronchioles in a dose-dependent manner.
Masson's trichrome staining revealed dense collagen deposition/sub-epithelial fibrosis throughout the interstitium surrounding the airways and vessels of lung tissues in the OVA group compared to the control group (Fig. 5A-D). PEDF treatment obviously reduced collagen deposition compared to the OVA group (P<0.05, Fig. 5E).
Total lung hydroxyproline content was significantly increased in the OVA group compared to the control mice (P<0.05, Fig. 5F). In contrast, treatment with PEDF resulted in a significant reduction in total lung hydroxyproline content compared to the OVA group.
Additionally, α-SMA immunostaining was used to evaluate the thickness of airway smooth muscle. A marked increase in the α-SMA-staining areas in the peribronchiolar region was found in the OVA mice compared with the control mice (Fig. 6A-D). Administration of PEDF significantly reduced the area of the α-SMA-stained smooth muscle layer in a dose-dependent manner compared to the OVA group (P<0.05, Fig. 6E).
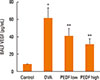
PEDF ameliorates the expression of VEGF protein in lung tissue and BALF in chronic experimental asthma
To determine whether VEGF is inhibited by PEDF in this model, we evaluated VEGF protein expression in mouse pulmonary tissue and VEGF levels in BALF. Compared to the control group, the OVA group had higher VEGF protein levels in the pulmonary tissue, and PEDF treatment prevented this increase (P<0.05, Fig. 7). ELISA also showed a marked increase in BALF VEGF levels in OVA-challenged mice. The mice treated with PEDF demonstrated lower levels of VEGF in BALF compared to the OVA group (Fig. 8). We also observed that PEDF administration reduced VEGF in a dose-dependent manner in the BALF and lung tissue of OVA-challenged mice.
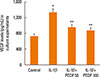
PEDF suppresses IL-1β-induced VEGF release in BEAS-2B cells
VEGF released from airway epithelial cells can aggravate airway inflammation and airway remodeling. To further ascertain the anti-inflammatory and antiremodeling mechanisms of PEDF, we studied the effects of PEDF on IL-1β-induced VEGF expression in BEAS-2B cells. The levels of VEGF in culture supernatants from BEAS-2B cells induced by IL-1β were significantly increased compared to unstimulated cells. PEDF evidently inhibited the release of VEGF from BEAS-2B cells stimulated with IL-1β in a dose-dependent manner (P<0.05, Fig. 9).
DISCUSSION
Airway remodeling is an important pathophysiological characteristic of chronic asthma and correlates with the duration and severity of asthma.8 In this study, we evaluated the effects of recombinant PEDF on airway remodeling in chronic allergic asthma. Our results showed that recombinant PEDF had significant inhibitory effects on eosinophilic airway inflammation, AHR, and airway remodeling in this asthma model. In addition, treatment with recombinant PEDF resulted in marked inhibitory effects on the high expression of VEGF in lung tissues of OVA-challenged mice. Furthermore, recombinant PEDF administration suppressed VEGF release from BEAS-2B cells stimulated with IL-1β.
Th2-mediated airway inflammation present within the lung is fundamental in asthma. The Th2 cytokines IL-4, IL-5, and IL-13 are considered the key molecular mechanisms of asthma and have been shown to be associated with airway inflammation.2021222324 In the present study, the production of Th2 cytokines was significantly increased in OVA-challenged mice. Treatment with PEDF inhibited the levels of Th2 cytokines, such as IL-4, IL-5, and IL-13 in BALF, compared to OVA-challenged mice. In addition, recombinant PEDF prevented an increase in total inflammatory cells, especially eosinophils. Lung histological analysis also demonstrated that recombinant PEDF suppressed inflammatory cell infiltration, mucus hypersecretion, and goblet cell metaplasia in the airway. The results indicated that recombinant PEDF exerts significant anti-inflammation effects in chronic OVA-induced allergic mice.
Collagen deposition/subepithelial fibrosis in the airways, an important aspect of airway remodeling, is related to clinically severe asthma and a decline in pulmonary function.25 Previous studies showed that PEDF possesses antifibrosis properties exerted via inhibition of the TGF-β1 expression.2627 TGF-β1, a pro-fibrotic cytokine, leads to subepithelial collagen and extracellular matrix protein deposition in asthma.28 Thus, a marked increase in collagen deposition was observed around the airway in chronic OVA-induced mice. We show for the first time that the administration of recombinant PEDF not only significantly inhibits the TGF-β1 level in BALF in ova-induced mice, but also prevents subepithelial collagen deposition, as determined by Masson's trichrome staining and hydroxyproline analyses in chronic allergic asthma.
Airway smooth muscle hyperplasia is another key characteristic of airway remodeling and appears to be attributable to exaggerated AHR in asthma.29 In this study, the α-SMA stained area was significantly increased in chronic OVA-exposed mice. The chronic OVA-induced increase in airway smooth muscle was significantly inhibited by recombinant PEDF administration. The ability of recombinant PEDF to suppress airway smooth muscle thickness may be an important reason for its suppression of AHR.
VEGF is a potent stimulator of angiogenesis and structural changes in asthma.30 VEGF can aggravate Th2-type airway inflammation via IL-13-dependent and -independent mechanisms.31 Epithelial cell-derived VEGF promotes airway remodeling in asthma.32 VEGF levels are increased in sputum and lung tissues from asthmatic patients and correlate with the disease severity of asthma.333435 The inhibition of VEGF can lead to a significant reduction in basement membrane thickness and goblet cell hyperplasia.36 Previous studies showed that PEDF counteracts the biological actions of VEGF via down-regulating VEGF expression or binding VEGF receptors.3738 The results of our study showed that expression of the VEGF protein was enhanced after chronic OVA challenge, and treatment with recombinant PEDF decreased VEGF protein expression in the lung tissues and VEGF levels in the BALF of chronic OVA-induced allergic mice. In addition, recombinant PEDF significantly suppressed VEGF release from BEAS-2B cells stimulated by IL-1β. Therefore, we hypothesized that recombinant PEDF may inhibit airway inflammation and airway remodeling by reducting the expression of VEGF in a chronic asthma model.
In conclusion, our data demonstrated that PEDF exerts significant anti-inflammatory and antiremodeling effects in a chronic allergic asthma model. In addition, recombinant PEDF obviously inhibited VEGF expression in vivo and in vitro. Our findings indicate that PEDF might be a therapeutic target for chronic allergic asthma.


 XML Download
XML Download