

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Allergy to plant-derived food is often associated with a primary sensitization to pollen allergens. A well-known and clinically important phenomenon is the "celery-mugwort-birch-spice syndrome", which occurs in adults12345 as well as in children.6 The additional relationship between mugwort and ragweed may possibly even extent this cross-reactivity syndrome.7 Sensitization against celery tuber represents one of the clinically most important causes of food allergies, especially in European countries such as Switzerland, Germany, and France.8910 Derived from the same botanical Apiaceae family, the cross-reactive spices anise, fennel, cumin, and coriander may complicate the diagnosis.1 In the celery-mugwort-birch-spice syndrome, IgE cross-reactivity is associated with several classes of allergens: (1) Bet v 1, Api g 1, and other PR-10 allergens, (2) profilins such as Bet v 2 or Api g 4,11 (3) non-specific lipid transfer proteins,1213 and (4) high molecular weight (HMW) allergens (32-70 kDa), of which only Api g 5 from celery and its homologues from grass pollen and fennel have been identified so far.1416 Sensitization to Api g 5 was detected in up to 42% of patients with celery allergy, and there was an excellent correlation between sensitization to Api g 5 and to cross-reactive N-glycan epitopes.1517 Monoclonal antibody, BIP3 was originally raised against birch pollen HMW allergens between 32 and 63 kDa.5 It also reacts to mugwort pollen and celery antigens at 55-58 kDa, which were previously characterized by N-terminal sequencing.1718 The resulting glycoproteins turned out to be size variants of Api g 5.19
In the present study, we aimed to generate peptide mimotopes using this cross-reactive antibody BIP3, in analogy to previous studies.20 Therefore, we screened a peptide phage display library with BIP3 and immunized BALB/c mice with the phagedisplayed BIP3-specific mimotopes. The immune sera were used to reveal reactivity towards glycoprotein Api g 5. Furthermore, we addressed the possible participation of carbohydrate moieties, i.e. N-glycans in the BIP3 epitope by testing for cross-reactivity to horseradish peroxidase (HRP) as an ideal model glycoprotein representing multiple carbohydrate species21: Whereas bromelain from pineapple stem has only one xylose- and fucose-containing N-linked glycan (MUXF), HRP has 6 xylose and fucose containing glycans (MMXF). This is important as Api g 5 has both types of fucosylated and xylosylated complex N-glycans, namely, MMXF and MUXF.17 It should be noted that MUXF from bromelain lacks a third mannose residue which is found in many plant glycoproteins like on HRP and Api g 5. This might influence, to some extent, antibody binding as bromelain has been found less sensitive for detection of CCD-positive sera than HRP and Api g 5. Our hypothesis was that mimotopes could be used for improved diagnosis and immunotherapy of the celery-mugwort-birch-spice syndrome.
MATERIALS AND METHODS
Monoclonal antibodies and patient sera
The production of mAbs BIP3 and BIP1 has been described by Jarolim et al.5 In that study, birch pollen extract was used for immunizing BALB/c mice. Among the monoclonals raised, some (e.g. BIP1 and BIP4) were directed against the major allergen Bet v 1, whereas only BIP3 was directed against HMW allergens. Sera from birch pollen allergic patients cross-reacting to celery were sampled in the Vienna outpatient clinic Allergy-Care®. The usage of serum remnants from routine diagnosis was permitted in written form by the patients, and the study was approved by the ethics committee of the Medical University of Vienna, according to the Helsinki guidelines.
Extracts and proteins
Birch and mugwort pollen were purchased from Allergon AB (Engelholm, Sweden) and were extracted (10% weight/volume) using 10 mM/L potassium phosphate buffer (pH 7.0) overnight under continuous agitation at 4℃. After 1 hour centrifugation at 4℃ (40,000 g), supernatants were dialyzed (cutoff 6,000-8,000 Da) against 10 mM/L K2HPO4 and 3 mM/L sodium azide for two days. Thereafter, dialysates were lyophilized and stored at -20℃. Apiaceae spices (anise; coriander; cumin; fennel) and celery tubers were purchased at a local store, ground in liquid nitrogen, and extracted as described above. Protein content of extracts was determined by Bradford assay.22 Api g 5 was purified as described by Bublin et al.17 HRP was purchased from Fluka, and the plasmid for the production of rPhl p 5 was kindly provided by Prof. A. Petersen, Research Center, Borstel, Germany.
SDS-PAGE, Western blots and deglycosylation
Protein extracts were loaded onto gels in the following concentrations: coriander 50 µg/lane, cumin 50 µg/lane, fennel 50 µg/lane, anise 50 µg/lane, mugwort pollen 50 µg/lane, celery 25 µg/lane, birch pollen 25 µg/lane, Api g 52 µg/lane, and HRP 4 µg/lane. They were separated by preparative 8% or 12% SDS-PAGE under reducing conditions. Separated proteins were transferred to activate PVDF membrane (GE Healthcare, Little Chalfont, Buckinghamshire, UK). Blot strips were saturated with blocking buffer 5% dry milk in Tris buffered saline (TBS) with 0.05% Tween-20 (TBST).
The mAb BIP3 and the Bet v 1-specific mAb BIP1 were diluted to 1 µg/mL in blocking buffer; the sera of mimotope-immunized mice (see below) were diluted 1:100 and incubated with blot strips at 4℃ overnight. After washing with TBST, bound mouse mAbs were detected by125 I-labelled sheep anti-mouse Ig (Amersham Life Science, Little Chalfont, UK) and blots autoradiographed on Kodak Biomax MS films at -70℃.
For human IgE detection, after saturation of blot strips, individual human sera diluted 1:5 in blocking buffer were incubated overnight. After washing with TBST, bound IgE was detected by a peroxidase-labeled anti-human IgE (diluted 1:3,000) (KPL, Gaithersburg, MD, USA). After washing, the reaction was developed with Clarity ECL Western substrate (BioRad, Contra Costa County, CA, USA), and data were acquired by using ChemiDoc Touch Imaging System (BioRad, CA, USA). Alternatively for competition assays, blots were first pre-incubated with a pool of 5 mouse mimotope sera and were incubated with a patient's serum 1:5 after stringent washing with TBST. IgE was detected as described.
Deglycosylation of blotted proteins was performed by 3 hours of incubation of nitrocellulose membranes in 0.01 M NaIO4 in acetate buffer (50 mM pH 4.5) at 4℃ under continuous shaking while protected from light. The reaction was terminated by washing in aqua bidest.
Biopanning
BIP3 specific phage clones were selected from a random peptide phage library expressing circular (pVIII9aa.Cys) nonapeptides fused to pVIII of the filamentous bacteriophage fd.23 Libraries were kindly provided by IRBM (Istituto di Recherche di Biologia Molecolare P. Angeletti SPA, Rome, Italy).
Three rounds of biopanning were performed using 1 µg of monoclonal antibody BIP3 in 0.1 M NaHCO3, pH 8.5 for coating on ELISA plates (Nunc, Roskilde, Denmark) overnight at 4℃ in a humid chamber. After blocking using PBS containing 3% BSA for 1 hour at 37℃, 1011 plaque-forming units (pfu) of phage library or eluent from the previous round of panning were added to each well and incubated at 37℃ for 2 hours. Unbound phages were removed by washing with TBS/0.5% Tween 20. BIP3-binding eluted using 0.1 M HCl pH 2.2 containing 1 mg/mL, and pH was adjusted to 7.0 with 1 M Tris-HCl, pH 8. These phage clones were used for further rounds of biopanning. The enrichment of BIP3-specific phage particles was confirmed by colony screening.
Colony screening
After each biopanning round, eluted phages were propagated in E. coli XL-1 Blue co-infected with helper phage VCSM13 (Stratagene, La Jolla, CA, USA) in liquid culture. An aliquot of phages was further cultivated on LB/carbenicillin plates, and the total number of eluted phages was calculated by counting colonies. Sixty colonies/biopanning round were randomly picked, transferred to new LB/carbenicillin plates, and grown overnight at 37℃. The colonies were then incubated for 1 day covered with an isopropyl-β-D-1-thiogalactopyranoside (IPTG)-treated nitrocellulose. After fixing the colonies to the nitrocellulose by incubation in a chloroform atmosphere and 30-minute blocking using 50 mM Tris, 150 mM NaCl, 5 mM MgCl2, and 3% (w/v) BSA, pH 8.0, cells were lysed using blocking buffer additionally containing 400 mg/mL lysozyme (Sigma, St. Louis, MO, USA) and 20 U/mL DNAse (Boehringer-Mannheim, Mannheim, Germany) for 1 hour at room temperature. Then the filters were saturated with PBS containing 0.5% BSA and 0.5% Tween 20 for 1 hour at room temperature. Afterwards, the membrane was screened for BIP3-specific phage clones using antibody BIP3 in comparison to BIP1 as isotype control antibody and detection performed as described in section "SDS-PAGE and Western blot." Positive clones were re-amplified in overnight cultures and stored in 20% glycerol at -70℃.
DNA purification and sequencing
BIP3-specific clones were lysed and DNA was isolated by precipitation with polyethylene glycol 8000 (Amresco, Dallas, TX, USA). DNA sequencing was performed by the Sanger dideoxy method using a Thermo Sequenase Cycle Sequencing Kit (Amersham Life Science) with fluorescence-labeled primer 5'-GCT TTA CAC TTT ATG CTT-3' (Eurofins, Ebersberg, Germany) using a LI-COR DNA sequencer 4000L (LI-COR Inc., Lincoln, NE, USA). Phylogenetic alignments were constructed using Vector NTI Advance 11.5.0 (Invitrogen, Carlsbad, CA, USA).
IgE inhibition experiments with mimotope peptides
Birch pollen allergic patients' sera were diluted 1:5 in blocking buffer or in blocking buffer containing 107 cfu/mL of single BIP3-mimotope clones from the third biopanning round. As negative controls, the original library, helper phage (VSCM13), or a phage clone of irrelevant specificity (CRQTRTRTMPGCG) (107 cfu each), were used. Diluted serum samples were then incubated on blotted birch pollen extract at 4℃ overnight and IgE detected as described above.
Immunization experiment
BALB/c mice (female; 3-5 weeks) were purchased from Charles River Labs, Sulzheim, Germany and were treated according to the European community rules of animal care. Immunizations were performed with the permission of the Austrian Ministry of Science (BMWF-66.009/0170-II/10b/2009), and institutional and national guidelines for the care and use of laboratory animals were followed.
Mice were immunized intraperitoneally (i.p.) with either of the following mimotope phage clones without adjuvants: 1-12-cyclo-CKASSCDTGHC; 1-12-cylo-CFFAWRSLPNC, and 1-6-cyclo-CHKLRCDKAIA on days 0, 14, and 28 using 1013 phage particles/100 µL 0.9% NaCl/PBS. Phage clone CHKLR CDKAIA was used for immunization of additional BALB/c mice (n=6) according to the same treatment scheme. Blood was taken on days 0 (pre-immune serum), 21, and 38.
Testing anti-BIP3 mimotope mouse sera in ELISA
Microtitre plates (Maxisorp, Nunc, Roskilde, Denmark) were coated overnight at 4℃ with 1 µg/mL of Api g 5, HRP, or for control, rPhl p 5 in 50 mM NaHCO3, pH 9.6 overnight at 4℃. Plates were washed with Tris-buffered saline containing 0.05% Tween 20 (TBST) and blocked for 2 hours at room temperature with TBST/1% bovine serum albumin (BSA). Sera from the immunisations with phage CHKLRCDKAIA were pooled to equal parts, diluted 1:100 in TBST/0.1% BSA, and incubated overnight at 4℃. Plates were washed, and alkaline phosphatase labeled anti-mouse IgG/M in TBST/0.1% BSA was added for 2 hours at room temperature. Reaction was developed with para-nitrophenylphosphate in 0.2 M/L Tris buffer. Optical density was measured at 405 nm using microplate reader Tecan infinite M200-Pro (Tecan Group Ltd., Männedorf, Switzerland).
For mouse IgG titer determination, plates were coated with 10 µg/ml Api g 5 and incubated with serial dilutions 1:10-1:10,00 of anti-BIP3 mimotope CHKLRCDKAIA mouse serum pool (5 sera pooled at equal parts) or pre-immune serum pool of the same mice. Bound IgG was detected with a 1:5,000 dilution of goat anti mouse IgG-Fc Fragment-HRP (Bethyl Laboratories Inc., Montgomery, TX, USA), ABTS was used for color development. Optical density was measured at 405 nm.
RESULTS
BIP3 recognizes cross-reactive birch, mugwort, and celery antigens
The monoclonal anti-birch pollen antibody BIP3 reacted to HMW allergens at 35-77 kDa in birch pollen; its binding was almost, but not completely, abolished when N-glycans were chemically removed (Fig. 1A).
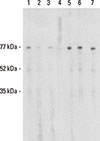
BIP3 also exhibited binding to HMW allergens in birch, celery, and mugwort pollen extracts (Fig. 1B), which is indicative of the cross-reactivity spectrum typically seen in sera of patients with the celery-mugwort-birch-spice syndrome. For comparison, lane 1 in Figure 1B shows varying binding intensity of the anti-Bet v 1 antibody BIP15 to the same extracts.
Supplemental Figure 1 shows the IgE-binding patterns of 3 different patients (A, B, and C) suffering from the celery-mugwort-birch-spice syndrome and tested on 1) the Apiaceae spices anise, fennel, coriander, and cumin, 2) mugwort pollen, celery, and 3) nApi g 5 and HRP. Note that patients A and B recognized more Api g 5 than HRP, whereas patient C showed the opposite, indicating that carbohydrate decorations of allergens are important for some, but not all patients.
Mimotopes as BIP3 epitope surrogates, generated by biopanning
To generate mimotopes for the HMW allergens, biopanning was performed using the monoclonal antibody BIP3 for screening. During biopanning rounds, the total titers of eluted phages increased continuously from 2.4×104 to 6.2×107 (Fig. 2A). In parallel the proportion of clones testing positive in colony screening assay increased from 14 to 44 out of 60 clones randomly picked after each round (Table). Phage clones that showed high binding capacity for BIP3 in colony screening were sequenced. The deduced amino acid sequences (Table) showed that in panning round 3, one clone (CHKLRCDKAIA) dominated, representing 6 out of the 8 sequenced clones. The 6 amino acid sequences were grouped in a phylogenetic tree according to similarity (Fig. 2B). The dominant clone CHKLRCDKAIA as well as 2 additional clones (CKASSCDTGHC and CFFAWRSLPNC) were chosen for further studies.
BIP3 mimotopes imitate a human IgE epitope and are immunogenic
To prove the mimicry potential of the selected phage clones with the original epitope of BIP3, inhibition studies were performed with human sera. Indeed, pre-incubation of a human birch-pollen specific serum with the 3 selected phage-displayed-mimotopes resulted in specific reduction in IgE binding to the HMW allergens. Incubation of IgE with an unrelated phage clone, the original phage library, or helper phage did not have any inhibitory effect (Fig. 3).
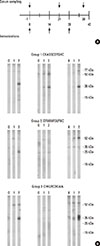
When BALB/c mice were immunized with the BIP3 mimotopes (Fig. 4A), all formed specific IgG to HMW birch pollen allergens (Fig. 4B), except in the mice of groups 2 and 3, as shown in the middle panels in Fig. 4B. The formed IgG had a pattern similar to that of BIP3 (Fig. 1A) or IgE from human patients tested on various cross-reactive extracts of relevance in the celery-mugwort-birch-spice syndrome (Fig. 3, lane 1; and Supplemental Figure 1).
Verification of Api g 5 as the BIP3 target using mimotope-induced antibodies
So far, we could show that the BIP3 mimotopes via molecular mimicry induced IgG against birch pollen HMW allergen. Consequently, we next aimed to reveal the molecular target of BIP3. We pooled sera of mice immunized with the dominant mimotope phage clone CHKLRCDKAIA and tested them for IgG binding to purified glycoprotein Api g 5 (of which no birch pollen homologue has been characterized so far), to HRP as a model glycoprotein, or to non-glycosylated control allergen rPhl p 5. IgG of BIP3-mimotope-immunized animals showed strong binding to Api g 5, moderate binding to HRP, and no reactivity to Phl p 5 (Fig. 5A). The induced IgG titers to Api g 5 reached at least 1:500 (Fig. 5B). When the sera of BIP3 mimotope-immunized mice were pre-incubated to blotted spices, mugwort pollen, celery, Api g 5, and HRP, they reduced IgE binding of serum of patient C which was subsequently incubated (Supplemental Figure 1, panel "C inhib"). The results indicate that Api g 5 and its cross-reactive homologues are the major targets of BIP3, with carbohydrates playing only a minor role in its epitope. Furthermore, phage-displayed BIP3 mimotopes were immunogenic and induced specific IgG titers to Api g 5.
DISCUSSION
Carbohydrate-specific IgE antibodies are mainly seen as complicating factors in allergy diagnosis as their clinical relevance is still debated. These IgE antibodies specific for cross-reactive carbohydrate determinants (CCDs) have a high prevalence as they are found in 15%-30% of allergic patients' sera.242526 Especially in the celery-birch-mugwort spice syndrome,12482728 HMW allergens decorated by carbohydrates, contributing to their resistance, have been characterized in the past.1025 In this clinical cross-reactivity syndrome, up to 50% of IgE is targeting the celery glycoprotein Api g 51719 of the berberine bridge enzyme-like protein family, of which so far only a grass pollen homologue Phl p 4,14 but none from birch or Apiaceae spices, has been cloned. Our in silico attempts to localize the epitope by structural alignments failed. In accordance with previous work,17 we could show in this study that BIP3 type antibodies react to HMW-allergens in these allergen sources. Our data also suggest that their targets are Api g 5 and its cross-reactive homologues, and that carbohydrates play a minor role in this binding.
Carbohydrate-specific IgE antibodies are prevalent in 15%-30% of allergic patients' sera.242526 In general, they represent a mixture of N-(asparagine)-linked oligosaccharides containing β-1,2-xylose and core α-1,3-fucose29 and belong to the most abundant environmental immune determinants. On the one hand, anti-CCD IgEs have been associated with false positive diagnostic results,30,31 but on the other hand, anaphylactic reactions associated with IgE specific for carbohydrate oligosaccharides, such as galactose-α-1,3-galactose (α-Gal), have been reported.323334 It has also been shown for tomato β-fructofuranosidase Sol a 12 (previously Lyc e 2) possessing multiple carbohydrate moieties, that the glycosylated, but not the deglycosylated protein was able to induce histamine release in mast cells passively sensitized with sera of tomato allergic CCD reactive patients.35 Notably, the affinity of anti-CCD IgE may reach a dissociation constant of 10-10 M/L.36
Our study hypothesized that the BIP3 mimotopes, as epitope surrogates independent of protein-, carbohydrate-, or mixed-nature of the original antigen, should allow bypassing of the uncertain aspects of carbohydrate moieties by translating the epitope into peptides. Active immunotherapy with peptide mimotopes may be of great advantage as they induce blocking IgG, but do not activate inflammatory allergen-specific T cells.373839 Indeed, the mouse immunization studies have demonstrated that the BIP3 mimotopes are immunogenic and that the induced IgG is directed towards allergens relevant for human IgE binding in the celery-birch-mugwort-spice syndrome. This result was supported by the fact that incubation with the mimotopes could substantially decrease IgE binding to the HMW birch pollen proteins.
In the celery-mugwort-birch-spice syndrome, celery has been recognized as a central allergen. The celery allergen Api g 5 consists of a mixture of 2 polypeptides with molecular weights of 53 and 57 kDa, carrying N-glycans of the MUXF3 type, resembling the HMW-allergens relevant in the clinical syndrome. The BIP3 mimotopes induced IgG binding to Api g 5, identifying this allergen as the major target of BIP3 and IgE specific for HMW. Carbohydrates contribute to the Api g 5 IgE epitope, and a recent study showed excellent correlation between IgE reactivity against Api g 5 and MUXF3 (Manα1-6(Xylβ1-2)Manβ1-4GlcNAcβ1-4(Fucα1-3)GlcNAc).40 However, the minimal binding of the induced mouse IgG to model glycoprotein HRP suggests that the BIP3 mimotopes predominantly mimic a protein epitope.
Taken together, we report here the generation of peptide mimetics of a cross-reactive Api g 5 epitope relevant in the celery-mugwort-birch-spice syndrome and propose them as candidates for epitope-specific immunotherapy within this clinical syndrome.



 XML Download
XML Download