

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Food allergy is an important medical and social problem and its incidence appears to be on the rise.1 Soy, which belongs to the "Big-8 allergenic foods",2 is widely used by the food industry, can be present as excipients in non-food products (gasolines, inks, crayons, adhesives, soaps, pharmaceuticals and cosmetics),3 and may act as an aeroallergen causing occupational allergy in rural workers and in individuals involved in soybean processing.4,5,6 Thus, it is difficult for sensitive individuals to avoid exposure to these ubiquitous proteins. However, the incidence of soy allergy is lower than generally expected.7,8,9,10
The identification and characterization of soy components responsible for allergy is essential for the development of immuno-intervention strategies. Three soybean proteins, Gly m Bd 30 K/P34, Gly m Bd 28 K/P28 and Gly m 5, have been described as major soy allergens based on IgE recognition.11,12,13,14,15 However, the in vivo allergenicity has not been investigated. Gly m Bd 30 K/P34 is an outlying member of the papain-superfamily of cysteine proteases.16,17,18 Despite a mutation in the active site that silences the protease activity,16 it has been characterized as a major allergen,11,12 and several linear B epitopes have been mapped.19,20 Nonetheless, its clinical relevance should be explored.
Considering the low prevalence of hypersensitivity to soybeans, it has been proposed that soy allergy may occur through secondary sensitization. Co-sensitization to soy has often been described in Central Europe in birch pollen-sensitized patients.21,22 We have previously shown that Gly m 6 G4 and Gly m 5 are recognized by the IgE antibodies of patients allergic to milk, although the clinical relevance of this finding requires further investigation.23,24,25 Taking into account the fact that soybean proteins are ubiquitous, exposure to soybean in patients is difficult to discard. A milk-allergic mouse model was used to overcome this problem, and we previously demonstrated that hypersensitivity reactions were elicited in milk-sensitized animals exposed to total soybean proteins or Gly m 6 G4. These findings showed the potential allergenicity of Gly m 6 G4 as a cross-reactive soy component.23,24,26
In this study, we investigated the recognition of Gly m Bd 30K/P34 by cow's milk protein (CMP)-specific antibodies and evaluated the clinical relevance of this cross-reactivity using the milk allergy mouse model. We found that P34, one of the main allergens of soy, behaves as a cross-reactive allergen with bovine caseins, which are considered one of the main allergens in cow's milk.27,28 These findings increase our understanding of the clinical intolerance observed in a restricted proportion of milk allergic patients (10%) treated with a soy-based formula.29 In addition, an allergen immunotherapy could be developed based on this biological phenomenon.
MATERIALS AND METHODS
Protein extracts and antibodies
Soybean protein (SP) extract was obtained from Glycine max L. Merr. seeds as described previously.24 Briefly, seeds were crushed and extracted with 0.01 N NaHCO3 at 90℃. The extract was centrifuged at 2,500×g for 20 minutes at room temperature and lipids were extracted with chloroform overnight at 4℃. The extract was dialyzed against distilled water and stored at -20℃ until use.
CMP extract was obtained from commercial skimmed milk. Proteins were extracted with phosphate-buffered saline pH 7.4 (10 mg/mL) and filtered. The extract was stored at -20℃ until use. The presence of soy components in the CMP extract was previously discarded by indirect ELISA with SP-specific rabbit antiserum.
Sera from 10 pediatric patients diagnosed with milk allergies according to history, skin prick test, and serum specific IgE were used. Double-blind placebo-controlled food challenge is not performed in Argentina for diagnosis; instead, elimination of milk from the diet and open challenge is performed. Soy allergy was ruled out based on history and serum IgE against SP. Sera from healthy individuals with no allergy history and normal level of serum IgE, or from patients allergic to aeroallergens with no CMP-specific IgE antibodies or history of food allergy, were included as negative controls.
Three monoclonal antibodies (mAb) with differential specificities for α-casein (1D5), β-casein (4C3) and κ-casein (3B5), which were characterized previously,30 were used.
Gly m Bd 30K/P34 construct expression and purification
The cDNA coding sequence for P34 (GenBank: DQ324851) was obtained by PCR amplification of the cDNA library.31 Amplified PCR products were cloned directionally into pENTR/D TOPO (Life Technology, S.A. Argentina) and were then transferred to the pDEST-maltose-binding protein (MBP) destination vector for expression.32 E. coli BL21 Codon Plus containing the constructs pDEST His-MBP-P34 and pDEST His-MBP were induced and recombinant proteins were purified as described previously.24 Depletion of lipopolysaccharide was performed with a Sepharose-polymyxin B resin (Pierce, Rockford, IL, USA). Endotoxin determination was performed using the Limulus amoebocyte chromogenic assay (LONZA, Buenos Aires, Argentina). Protein concentration was determined using the bicinchoninic acid assay with bovine serum albumin as a standard (Pierce, Rockford, IL, USA).
Immunochemical assays
Immunoblotting
Purified P34 and MBP (3 µg) were separated by SDS-PAGE and transferred to nitrocellulose membranes. Blocked membranes (3% horse serum) were incubated with patient serum (1:5, overnight at 4℃), followed by biotinylated anti-human IgE monoclonal antibody (1:3,000, 4 hours at 37℃, Vector Laboratories Inc., CA, USA), and finally, with horseradish peroxidase (HRP) -streptavidin conjugate (1:3,000, 30 minutes at 37℃, Sigma-Aldrich, MO, USA). Luminol chemiluminescent substrate and exposure to X-ray film (Amersham Hyperfilm ECL, GE Healthcare Bio-Sciences Corp., USA) were used for band visualization. Membranes were also revealed using the mAbs (1 µg/mL) followed by mouse immunoglobulin G-specific rat monoclonal antibody HRP-conjugate (1:3,000, 1 hour at 37℃ Bio-Rad Laboratories, CA, USA).
Competitive ELISA
Polystyrene microtitre plates were coated with 0.25 µg/well α-casein (Sigma-Aldrich, MO, USA) and blocked with 5% horse serum. Purified 1D5 mAb was separately incubated with various quantities of the soluble inhibitor α-casein, P34 or ovalbumin (OVA) as a non-related protein (2 hours at 37℃). This pre-mixed dilution was then added to the coated wells (30 minutes at 37℃). Finally, the secondary antibody was added (1:3,000, 1 hour at 37℃) and color was developed with o-phenylenediamine. Optical density (OD) was measured at 492 nm.
Flow cytometry-based basophil activation test (BAT)
Basophils were obtained from whole heparinized blood collected from non-allergic human donors using ammonium-chloride-potassium (ACK) buffer (0.15 M NH4Cl, 10 mM KHCO3, 0.1 mM Na2EDTA, pH 7.2) as erythrocyte lysis buffer. Total cells were re-suspended in ice-cold lactic acid buffer pH 3.9 (13.4 mM lactic acid, 140 mM NaCl and 5 mM KCl) for 5 minutes for stripping. Cells were washed and re-suspended again in 2 mL of saline buffer containing 20% serum from milk allergic patients (5 sera), 4 mM Na2EDTA and 10 mg/mL heparin (90 minutes at 37℃). Cells were washed with 20 nM HEPES buffer pH 7.4 and incubated with HEPES buffer containing 1 mM CaCl2 (30 minutes at 37℃).33 Cells were then challenged with whole milk proteins, SP or the recombinant P34 soy allergen at various concentrations. Saline buffer was used to stop cell activation. Finally, washed cells were incubated with 7-aminoactinomycin (7-AAD) (BD Pharmingen, USA) and biotinylated anti-human IgE monoclonal antibody (Vector Laboratories Inc., CA, USA), streptavidin-allophycocyanin (APC) (eBioscience, USA), phycoerythrin (PE)-conjugated antibody specific to human CD63 (BD Pharmingen, USA), or monoclonal antibodies specific to human CD203c (Macs, Miltenyi Biotec, Germany), followed by goat anti-mouse IgG fluorescein isothiocyanate (FITC) (Santa Cruz Biotechnology, USA). Fluorescence data were acquired with a Becton Dickinson FACSCalibur flow cytometer (Franklin Lakes, NJ, USA) and analyzed with the BD CellQuest Pro software and the FlowJo software (Tree Star Inc., Ashland, OR, USA). Controls with a non-related protein (OVA) or with sera from non-allergic subjects were included.
Sensitization and immune response to antigens
Mice sensitization and challenges
Male 6- to 8-week-old Balb/c mice were purchased from the School of Animal Sciences, University of La Plata, and kept under pathogen-free conditions with water and a commercial diet provided ad libitum. Mice were grouped into sensitization and control groups (n=8 per group). Animals were sensitized essentially as described previously,26 Ten days following the last boost, some mice were challenged with 20 mg of CMP or 5 mg of SP intragastrically (i.g.), while others were challenged with 5 µg of P34, 5 µg of β-lactoglobulin (β-Lg) (positive control antigen), or 10 µg of OVA (unrelated control antigen) by sublingual (s.l.) administration.
In vitro evaluation of the allergic reaction
Serum specific IgE antibodies were assessed by EAST using CNBr-activated cellulose paper discs coupled with CMP (1.75 mg/mL), SP (0.5 mg/mL), P34 (0.2 mg/mL) or MBP (0.1 mg/mL), and developed essentially as described previously.26 In addition, serum specific IgG1 and IgG2a were measured (ELISA) using CMP, SP, or P34 at 1 µg/100 µL, or MBP at 0.5 µg/100 µL, as described previously.26 Plasma histamine level was measured using competitive ELISA as described previously.26
To examine T cell activation, cytokines were measured in supernatants of antigen-stimulated spleen cells. Briefly, 24 hours following oral challenge, mice were killed, spleens were resected, and spleen cells were stimulated for 72 hours with CMP (350 µg/mL), SP (200 µg/mL), P34 (15 µg/mL) or MBP (7.5 µg/mL). Production of IL-5, IFN-γ (Invitrogen Corporation, USA) and IL-13 (R&D Systems, UK) was assayed by ELISA following the manufacturer's instructions.
In vivo evaluation of the allergic reaction
Symptoms were observed 30-60 minutes following oral challenge in a blinded fashion by 2 independent investigators, and were scored according to Table 1. Cutaneous tests were performed as described previously.26 Briefly, shaved skin was subcutaneously (s.c.) injected with 200 µg of CMP, 400 µg of SP, or 10 µg of P34 in 50 µL of sterile saline in one flank and with 10 µg of MBP or saline in the contra-lateral flank (negative control). Evans blue dye was intravenously (i.v.) injected and observation of a blue color in the first 30 minutes after injection was considered a positive cutaneous test.
Ethical considerations
The research was conducted in strict agreement with the international ethical standards for animal experimentation (Helsinki Declaration and its amendments, Amsterdam Protocol of welfare and animal protection and National Institutes of Health, USA NIH, guidelines: Guide for the Care and Use of Laboratory Animals) and was approved by the local Institutional Animal Care and Use Committee at the School of Animal Science (University of La Plata).
RESULTS
P34 is in vitro recognized by CMP-specific immunoglobulins
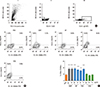
Human sera containing specific IgE antibodies were used to assess the recognition of recombinant P34. Fig. 1A shows that P34 was recognized as a coated antigen by 9 of 10 sera from milk allergic patients. Additional bands due to partial degradation of P34 were detected with some sera. No binding of MBP with sera from allergic patients (#3) or of P34 with sera from non-allergic subjects and non-milk-allergic patients (#11 and 12) was detected. To rule out co-sensitization in patients, immunoblotting was performed using three casein-specific monoclonal antibodies; all mAbs revealed a band corresponding to P34 (Fig. 1B). MBP was not recognized.
To rule out the possibility of artifactual reactivity due to new epitopes created during antigen coating, P34 was used as a soluble inhibitor in a competitive ELISA using the α-casein-specific 1D5 mAb (Fig. 1C). The sigmoid-shape of the dose-response inhibition curve obtained with P34 as an inhibitor supported the specificity of the antigen-antibody reaction. Antibody binding to the immobilized α-casein (0.25 µg/well) was inhibited by 100% with 0.002 mg/mL of soluble α-casein, while 50% inhibition (IC50) was achieved with 0.0002 mg/mL of α-casein and 0.05 mg/mL of P34. No inhibition was observed over a wide range of OVA concentrations.
To confirm the cross-reactivity with human IgE antibodies, we performed BAT using sera from milk allergic patients. Stripped basophils from healthy donors were passively sensitized with individual sera containing IgE and then activated with various concentrations of milk or soy proteins. A minimum of 100,000 events per sample were analyzed by flow cytometry and cells were gated based on physical properties (Fig. 2AI), 7 AAD-live cells (Fig. 2AII), and IgE+ cells (Fig. 2AIII), and were further analyzed for membrane CD63 and CD203c. Isotype controls were used to establish fluorescence thresholds. Percentages of double-positive cells were compared between basophils incubated with the allergen or PBS, CMP, and SP induced a higher percentage of IgE+ CD63+ CD203c+ cells compared with PBS alone (18.45%±1.35% and 18.80%±0.5% vs 9.16%±1.26%, respectively) (Fig. 2B). When P34 was used for IgE-dependent basophil activation, we observed 18.5%±0.45% double-positive cells. The non-related protein OVA at various concentrations resulted in 8.37%±0.31% double-positive cells. In addition, there were no statistically significant differences in cell activation using sera from non-allergic patients (data not shown).
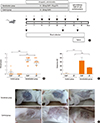
Total soybean proteins trigger acute allergic skin reactions in mice orally sensitized with CMP
CMP-sensitized Balb/c mice were used to investigate the clinical relevance of the immunochemical co-recognition (Fig. 3A). Intragastically sensitized mice developed hypersensitivity reactions (Fig. 3B) that were scored immediately after oral challenge with CMP or SP (Table 1). CMP-sensitized animals showed high scores upon challenge with SP, while no symptoms were observed in sham mice. To correlate these clinical findings with a systemic parameter, histamine was measured (Fig. 3C). We observed increased plasma levels in sensitized mice, demonstrating that the administration of SP or CMP to CMP-sensitized animals developed adverse reactions. Next, we examined the functionality of IgE antibodies. Subcutaneous injection of CMP or SP resulted in a local and distant inflammation within minutes (Fig. 3D). Injection of saline induced negative results in sham and sensitized animals.
P34 is recognized by serum antibodies and spleen cells from milk allergic mice
We next explored whether soy proteins were recognized by the secreted T-helper 2 (Th2)-associated antibodies. We found that SP and P34 were recognized by mouse CMP-specific IgE and IgG1 antibodies. No IgG2a specific antibodies were detected in sensitized or sham animals (Fig. 4A). To correlate the humoral immune response with the cellular immunity, the antigen-specific T-cell response and cytokine production were examined. A significant increase in the secretion of Th2 cytokines (IL-5 and IL-13) was detected in sensitized mice challenged with CMP or SP, while secretion of IFN-γ remained unchanged (Fig. 4B). Upon stimulation of spleen cells with detoxified P34 protein, IL-5 and IL-13 were secreted; in contrast, MBP did not induce cell stimulation.
P34 induces a positive cutaneous reaction and elicits immediate hypersensitivity symptoms
Consistent with the in vitro findings, we observed a positive cutaneous test with P34 (Fig. 4C), demonstrating that this molecule has at least two B epitopes capable of cross-linking membrane-bound IgE. Furthermore, the negative results obtained with all proteins in sham mice demonstrated that oral challenge with recombinant protein did not sensitize the mice.
Since sensitization does not always correlate with the clinical situation, a sublingual challenge was performed with P34 and control proteins in sham and sensitized animals. P34 and β-Lg strongly induced immediate symptoms in CMP-sensitized mice (Fig. 4D). In contrast, no clinical signs were induced by non-related OVA.
DISCUSSION
Although the population is highly exposed to soy allergens, exclusive soy allergy is rare.7 Instead, soy allergy is mainly encountered in patients primarily sensitized to other allergens due to cross-recognition of common epitopes.7 Despite the wide use of soybean formulae for infant feeding,34 there are few indications in atopic children. Allergy to soy proteins has been described in a small proportion of patients,35 and a randomized trial demonstrated that soy formula feeding does not induce an allergic reaction in milk-allergic children. Furthermore, some groups documented a clinical intolerance to soy-based formula in a small proportion of IgE-mediated milk allergic patients (<15%) that received soy products as a diary substitute.36 The American Academy of Pediatrics (AAP) and ESPGHAN recommended the use of extensively hydrolyzed CMP or a free amino acid-based formula as a first-line therapy if extensive hydrolyzed formula is not tolerated during CMA treatment. However, soy and rice protein-based infant formula are appropriate alternatives for these patients. ESPGHAN, AAP and DRACMA consensus recommended that soy formula should not be used in children (healthy or with food allergy) during the first 6 months of life as an initial treatment because of nutritional risks. In Argentina, soybean based-formula are commonly used as a dairy substitute for economic and acceptability reasons in children older than 6 months, and allergists are facing the problem that some CMA patients do not tolerate soy-based formula.37 The majority of these patients have serum IgE that recognizes SP; however, co-sensitization cannot be ruled out.37
We propose that the intolerance observed in a small number of children may be due to cross-reactivity between soy proteins and bovine caseins. We identified common structural determinants between caseins and two soybean proteins, Gly m 5 alpha subunit and Gly m 6 G4,23,24,25 and explored this immunochemical cross-reactivity using the IgE-mediated mouse model of CMA.26 In this study, we provide in vivo and in vitro experimental data that confirm the cross reactivity among non-phylogenetically related proteins, focusing on the major soybean allergen: Gly m Bd 30 k or P34.
Although the in vitro allergenicity of P34, including B and T epitopes,38 has been thoroughly described, its clinical relevance requires further study. In this report, we show that P34 was recognized by serum IgE of allergic patients, different casein-specific monoclonal antibodies, and CMP-specific IgE and IgG1 antibodies from milk-allergic mice. Consistent with the Th2-biased immune response, an immediate cutaneous reaction and hypersensitivity symptoms were observed in sensitized mice following oral exposure to P34. These findings are suggestive of at least two IgE epitopes with a surface distribution that enabled functional synapses between the allergenic molecule and the sensitized cells in different mucosa.
Unlike other cysteine proteases from tree and grass pollens, which have been associated with disruption of the protease-antiprotease balance at mucosal tissues,39 the allergenicity of P34 could be due to other physicochemical characteristics and not to protease activity. Here, we have shown using a variety of approaches that P34 contains B and T cross-reactive epitopes with CMP. This recombinant allergen promoted hypersensitive symptoms following sublingual challenge and induced a positive skin test in milk-sensitized mice. These findings demonstrate that the immunochemical cross-reactivity described for P34 may be clinically relevant. Further studies are required to characterize the differential immune response observed with the cross-reactive soy allergens and to delineate the scope and limitation of serum cross-reactive IgE. In this sense, animal models facilitate the investigation of the biological significance of this immunochemical cross-reactivity.
In conclusion, we identified P34 as a new cross-reactive soy protein that triggers hypersensitivity reactions in an IgE-mediated milk-allergic mouse model. This cross-recognition may explain the cross-allergenicity observed in some milk-allergic patients.



 XML Download
XML Download