

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Increasing evidence shows that exposure to indoor allergens is a causative factor for the development of allergic asthma among persons who are genetically predisposed to IgE autoantibody responses.123 Hypersensitivity to house dust mite (HDM) allergens is one of the most common allergic responses, and more than 80% of all patients with allergic asthma are sensitized to HDMs. These patients are characterized by high levels of serum total IgE and allergen-specific IgE to HDMs in Taiwan.456
Dermatophagoides pteronyssinus group 2 allergen (Der p 2) is considered a major allergen of HDM, and 80%-90% of all HDM-allergic subjects have been reported to be sensitive to Der p 2.4 Der p 2 allergens are proteins that tend to bind to the lymphocyte antigen 98 protein, also known as myeloid differentiation-2 (MD-2), which is associated with Toll-like receptor 4 (TLR4) on respiratory epithelial cells.7 MD-2 is an essential component of lipopolysaccharide (LPS) sensing, which upon binding to LPS and TLR4 forms the LPS-MD-2-TLR4 complex that can trigger cellular responses causing inflammatory reactions.78 Der p 2 has structural homology with MD-2, and this allergen has been reported to facilitate signaling through direct interactions with TLR4 in the absence of MD-2.8 It has been reported that mutations of the MD-2 gene on chromosome 8 can affect the human innate response.9 Since our previous reports demonstrated that mite allergen Der p 2 can trigger human B-lymphocyte activation and MD-2/TLR4 induction,10 it is conceivable that Der p2-susceptible genes may be related to the expression of MD-2.
It has been demonstrated that promoter polymorphisms can affect transcription activity and cell surface receptor expression.11 In our previous report, 6 SNPs of MD-2 promoter were identified and differences in genotype and allelic frequency were compared between allergic and healthy subjects; the results showed that the test genotypes of the rs1809441 and rs1809442 were significantly different between the 2 populations (P<0.05).12 MD-2 promoter SNPs (rs1809441/rs1809442) were significantly associated with Der p 2-specific IgE, suggesting that the 2 SNPs may play a major role in susceptibility to Der p 2- triggered immune responses in a Taiwanese population.12 Based on the 1,000 genome project data, the minor allele frequencies of the rs1809441 and rs1809442 are 0.467 and 0.474, respectively. The correlation of pairwise linkage disequilibrium (LD) between the 2 SNPs is D'=1.
The aims of this study were to examine whether promoter polymorphisms in the MD-2 gene can affect MD-2 expression and to clarify the relationship between Der p 2 allergy and SNPs of MD-2 promoter. Functional studies were conducted to investigate the role of MD-2 promoter polymorphisms in the development of allergic inflammation by analyzing the expressions of inflammatory cytokines in peripheral blood mononuclear cells (PBMCs) derived from allergic subjects with different genotypes of MD-2 promoter SNPs.
MATERIALS AND METHODS
Study subjects
The Institutional Review Board of Taichung Veterans General Hospital reviewed and approved the ethical issues of this study (IRB Nos. CF12009 and CF12010). Written informed consent was obtained from each participant before they were enrolled in the study. A total of 73 allergic patients who attended the Allergy Clinic at Taichung-Veterans General Hospital were recruited. There were 44 males and 29 females in the study. Their age ranged from 6 to 52 years, with a mean±standard deviation of 34±16. Among the 73 subjects, 45 suffered from allergic rhinitis (nasal symptoms), 38 suffered from atopic dermatitis or urticaria (skin symptoms), and 22 subjects suffered from asthma (airway symptoms); 32 had all the diseases. These subjects had a history of recurrent nasal stuffiness, nasal itching, sneezing, rhinorrhea, and/or asthma as defined according to the allergic rhinitis and its impact on asthma (ARIA) guidelines.1415 Serum total IgE and mite-specific IgE levels were measured using a UniCAP system (ThermoFisher Scientific, Uppsala, Sweden).
SNP genotyping analysis
Ethylene diamine tetra-acetic acid syringes with 5 mLof blood samples were collected, and DNA from the buffy coat was purified using a Genomic DNA Mini kit (Geneaid, Taoyuan, Taiwan). A set of primers were designed for the amplification of 156 base pair PCR products based on MD-2 promoter. The forward and reverse primer sequences were 5'-TggAgTgTAgTggCCCAATC-3' and 5'-CATggTgAAATTCCgTCTCT-3', respectively. The DNA of the 156 base pair PCR products was cleaned using a PCR DNA Fragments Extraction Kit (Geneaid). The genotypes were determined using a TaqMan real-time PCR (TaqMan SNP Genotyping Assays; Life Technologies, Carlsbad, CA, USA) in a StepOnePlus™ Real-Time PCR System (Applied Biosystems, Foster, CA, USA). Genotyping of 2 SNPs (rs1809441 and rs1809442) I n MD-2 promoter was performed using Custom TaqMan® SNP Genotyping Assay Mix (Rack ID: 4620713-1 and ID: 186819793-1) and TaqMan® Universal PCR Master Mix. The Taqman probe sequences for SNP genotyping of rs1809441 and rs1809442 in MD-2 promoter were AGCCTCCC(G/T)AGTAGC and CCTGGGTT(C/G)AAGCGAT, respectively. Two SNPs were identified at -1265 (rs1809441 G/T) and -1292 (rs1809442 C/G) in MD-2 promoter.
Transfection and luciferase reporter assay
The potential effects of different SNPs on promoter activity were investigated using a luciferase reporter gene assay system. The MD-2 promoter regions from different patterns of genotypes from subjects were constructed in the luciferase reported vector-pGL3 vector (Promega, Madison, Wisconsin, USA). A total of 5 plasmid constructs were prepared, which contained a promoterless (pGL3-basic) or wild genotype of CC (rs1809442)/GG (rs1809441), a single mutant genotype of CG and GT, a combined mutant genotype of GG and TT, or a positive control of SV-40 promoter. The epithelial cell line was transfected with 2 µg of construct plasmids (BEAS-2B) using FuGENE6 reagent (Roche Molecular Biochemicals, Indianapolis, IN, USA). At 24 hours post-transfection, the cells were treated with Der p 2 (10 µg/mL) for 24 hours. Luciferase activities were measured with a Luminoskan Ascent luminometer (Thermo Labsystems, Helsinki, Finland). Results were expressed as the fold increase in relative luciferase activity (RLA) of the MD-2 promoter construct vectors compared to the RLA of pGL3-Basic.
PBMC culture and cytokine analysis
The selection criteria of experiments for human PBMC culture and cytokine analysis were according to the written informed consent and the amount of specimen we obtained from each participant. A total of 73 allergic subjects who attended the allergy and clinical immunology outpatient clinics at Taichung Veterans General Hospital were recruited for the human PBMCs that used for culture. Human PBMCs were prepared by density centrifugation (Ficoll-Paque). The cells were stimulated with or without Der p 2 (10 µg/mL) or LPS (100 ng/mL) for 3 days in RPMI-1640 medium containing 10% heat-inactivated fetal blood serum (FBS) and 1% streptomycin/penicillin in a humidified 5% CO2 atmosphere. Cell culture supernatants were collected and evaluated for cytokine concentrations using commercially available Luminex MAP® kits according to the manufacturer's directions. Correlations between data sets were evaluated using Pearson's correlation coefficient (r).
Reverse transcription-polymerase chain reaction (RT-PCR)
PBMCs were cultured with or without Der p 2 or LPS (Escherichia coli strain 055:B5; Sigma-Aldrich, St. Louis, MO, USA) for 3 days, followed by RNA extraction. Total RNA from the cells was extracted using an RNeasy mini kit (Qiagen, Hilden, Germany). The first standard cDNA was synthesized by RevertAid M-MuLV reverse transcriptase (Thermo Fisher Scientific, Uppsala, Sweden) according to the manufacturer's protocol. The cDNA then served as a template in a PCR using a G-Storm PCR system. The forward and reverse primers were as follows: MD-2, 5'-AgAAgCAgTATTgggTCTgC-3' and 5'-ggCTCCCAgAAATAgCTTC-3'; GATA-3, 5'-CTCTgCTTCATggATCCCTAC-3' and 5'-CACAgTTCACACACTCCCTg-3'; Cε, 5'-CgTCTTCCCCTTgACCCgCTgCTg-3' and 5'-CACgTCCATgACCTgCCCgTCCTC-3'; Iγ1/2-Cµ, 5'-gggCTTCCAAgCCAACAgggCAggACA-3' and 5'-gTTgCCgTTggggTgCTggAC-3'; IL-1β, 5'-AAACAgATgAAgTgCTCCTTCCAgg-3' and 5'-TggAgAACACCACTTgTTgCTCCA-3'; IL-8, 5'-TTggCAgCCTTCCTgATTTCT-3' and 5'-TCTC AgCCCTCTTCAAAAACTTCTC-3'; and GAPDH, 5'-CCACCCATggCAAATTCCATggCA-3' and 5'-TCTAgACggCAggTCAggTCCACC-3'. The amplification cycles were as follows: 94℃ for 30 seconds, 60℃ for 30 seconds, and 72℃ for 60 seconds. The PCR products were then subjected to electrophoresis on a 2% agarose gel for 30 cycles. The electrophoresis products were visualized by ethidium bromide staining. The mRNA of GAPDH was used to control the sample integrity and loading.
Electrophoretic mobility shift assay (EMSA)
The electrophoretic mobility shift assay was performed according to the manufacturer's instructions as described in a LightShift® Chemiluminescent EMSA kit (Thermo Fisher Scientific Inc. Manufacturer: Pierce Biolechnology, Rockford, Illinois, USA). The oligonucleotide with the tandem GATA motif (-136 to -161) as a probe was 5'-CTCCGTATTTGATAAGGAACAAATAG-3'. The DNA-protein complexes were resolved on a 6% non-denaturing polyacrylamide gel and visualized by exposure to autoradiographic films.
PBMC culture, immunofluorescence staining, and flow cytometry
Cytokines in activated helper T (TH) cells derived from the donor's leukocytes underwent immunofluorescence staining and flow cytometry. Three-color staining methods were used to analyze the expression of IL-4 and interferon (IFN)-γ in CD4+ cells. PBMCs from 50-mL peripheral blood of allergic subjects were collected and stimulated with phorbol myristate acetate (PMA; 50 ng/mL), ionomycin (2 µM), and GolgiStop (Cytofix/Cytoperm Plus, Becton Dickinson: BD Pharmingen, Piscataway, NJ, USA) for 5 hours before washing twice with PBS. The cells were stained with peridinin chlorophyll-a protein (PerCP)-conjugated rat anti-human CD4 monoclonal antibody (BD Biosciences, Bedford, MA, USA) at room temperature for 30 minutes and washed with PBS. Cells were fixed with cytofix/cytoperm at room temperature for 30 minutes, washed with PBS, and stained with fluorescein isothiocyanate-conjugated rat anti-human IFN-γ monoclonal antibody (BD Biosciences) and R-phycoerythrin-conjugated rat anti-human IL-4 monoclonal antibody (BD Biosciences) at room temperature for 30 minutes and washed with PBS. The cells were resuspended in 0.5 mL of PBS with 0.1% (wt/vol) sodium azide. Mean fluorescence was measured using a flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA). A total of 5,000 cells were analyzed in each sample.
Statistical analysis
All statistical analyses were performed using SPSS software (version 22; SPSS Inc, Chicago, IL, USA). SPSS Sample Power 2.0 was used for power calculation analysis. The values are presented as mean±SEM for each group. Non-parametric analysis of the Wilcoxon signed rank and Mann-Whitney U tests were performed to calculate differences between the individual groups. A P value of less than 0.05 was considered statistically significant.
RESULTS
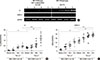
MD-2 promoter rs1809441(G/T) and rs1809442(C/G) SNPs affect transcription activity
The transcription activity on the MD-2 promoter of SNP No. rs1809441/rs1809442 was measured using a promoter assay. The effect of MD-2 promoter polymorphism transcription activity was analyzed using the transient expression system of a luciferase activity assay without Der p 2 stimulation. The schematic diagram of plasmid construct is shown in Fig. 1A. The promoter activities of several MD-2 promoter genotypes were different, which was significantly higher in MD-2 promoter SNPs (rs1809441-T/rs1809442-G) than in those with the genotype of (rs1809441-G/rs1809442-C) (P<0.01) (Fig. 1B).
The SNPs (rs1809441/rs1809442) of MD-2 promoter were searched for the transcription factor-binding site of Trans-acting T-cell-specific transcription factor-3 (GATA-3), based on the TFMATRIX transcription factor-binding site profile database TFSEARCH version 1.3.16 The sequence fragments of 71 bases were queried with 31 bases of rs1809441/rs1809442. The results showed that sequence fragments of rs1809441 highly correlated with GATA-3, with a score of 91.6. A similarly high correlation score was observed for rs1809442 with GATA-3 (Fig. 1C).
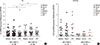
MD-2 promoter SNPs affect mRNA expressions of Cε and GATA-3
To determine whether the MD-2 promoter SNPs (rs1809441/rs1809442) affect Cε and GATA-3 mRNA expressions, PBMCs derived from allergic subjects were cultured with or without 24, 48, and 72 hours of Der p 2 challenge, followed by mRNA measurement, RT-PCR, and gel analysis presented by representative data (Fig. 2A). When mRNA expressions of Cε and GATA-3 were analyzed, both mRNA expressions could be up-regulated by Der p 2 in both groups of subjects. There were significantly increased expressions of Cε mRNA (Fig. 2B) and GATA-3 (Fig. 2C) with the Der p 2 challenge after 48-72 hours. The expressions of Cε and GATA-3 were significantly higher in allergic subjects with MD-2 promoter SNPs (rs1809441-T/rs1809442-G) compared to MD-2 wild-type (rs1809441-G/rs1809442-C) both in the conditions of PBMC cultured with or without Der p 2 stimulation after 48-72 hours (Fig. 2B and C).
MD-2 promoter rs1809441(G/T) and rs1809442(C/G) SNPs affect mRNA expressions of MD-2, Cε, Iγ1/2-Cµ, IL-8, and IL-1β
To determine whether the MD-2 promoter SNPs (rs1809441/rs1809442) affect MD-2 mRNA expression, PBMCs derived from allergic subjects were cultured with or without Der p 2, followed by mRNA measurement. There was a significant increase in MD-2 mRNA expression in subjects with MD-2 promoter SNP mutants, and the increase could be further up-regulated by Der p 2 (Fig. 3).
When mRNA and mRNA expressions of Cε and Iγ1/2-Cµ were measured, there was a significantly increased expression of Cε mRNA (Fig. 4A) and a significantly decreased expression of Iγ1/2-Cµ mRNA expression (Fig. 4B). These expressions were not affected by Der p 2 in any of the subjects with or without MD-2 promoter SNPs (rs1809441/rs1809442) (Fig. 4).
When the mRNA expressions of IL-1β and IL-8 were measured, the expression of IL-8 was higher in subjects with MD-2 promoter SNP mutations compared to the wild type, and both IL-1β and IL-8 could be up-regulated by Der p 2 in both groups of subjects. In comparison to the wild type, there were significantly higher expression levels in subjects with MD-2 promoter SNPs (rs1809441/rs1809442) after Der p 2 stimulation, although both IL-1β and IL-8 were up-regulated by Der p 2 (Fig. 5).
MD-2 promoter rs1809441(G/T) and rs1809442(C/G) SNPs affect cytokine secretion
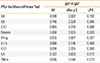
Next, we determined whether MD-2 promoter SNPs (rs 1809441/rs1809442) affect cytokine secretion. A total of 18 subjects were included in this study; 10 of whom had MD-2 promoter SNPs, but 8 did not. PBMCs were cultured with or without Der p 2 and LPS for 3 days, and the supernatants were collected for cytokine measurement using multiplex Enzyme-Linked Immunosorbent assays (ELISA). The results showed that there was a significantly higher level of IL-8 in subjects with MD-2 promoter SNPs (rs1809441/rs1809442) without stimulation. The levels of IL-1β, IL-6, IL-10, and TNF-α were up-regulated after Der p 2 stimulation in subjects with MD-2 promoter SNPs (rs1809441/rs1809442) (Table 1).
When cytokine secretion was compared between subjects with and without MD-2 promoter SNPs (rs1809441/rs1809442), the levels of IL-6, IL-8 and IL-10 were significantly higher in subjects with MD-2 promoter SNPs by Der p 2 stimulation. PBMCs stimulated with LPS showed a significant increase in IFN-γ and decrease in IL-4 in subjects with MD-2 promoter SNP mutations (Table 2).
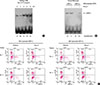
MD-2 promoter SNPs (rs1809441/rs1809442) affect the binding affinity of GATA-3 and TH2 cytokine expression
In order to clarify whether and how MD-2 promoter SNPs affect the binding affinity of GATA-3, and then affect the MD-2 promoter activity, the human promyelocytic leukemia cell line (HL-60) and human monocytes derived from donor' PBMCs (allergic and non-allergic subjects) were employed. The binding activity of GATA-3 to these MD-2 promoter SNPs was analyzed by EMSA using an oligonucleotide containing the tandem GATA motif (-136 to -161). The functional contribution of interactions between GATA-3 and allergen Der p 2 to MD-2 promoter activation, the binding complex was detected with the GATA motif (-136 to -161) probe. The binding activity of transcription factor-GATA-3 induced by Der p 2 (1.5 µg/mL) was in a time-dependent manner (0-8 hours) with the human promyelocytic leukemia cell line HL-60 (Fig. 6A), suggesting that there was a stimulatory effect of Der p 2 on the binding activity of GATA-3 after 4 hours. Furthermore, the monocytes derived from subjects with or without MD-2 promoter SNPs were collected to analyze differences in responses to Der p 2 between different genotypes. There was an obviously enhanced binding affinity of GATA-3 in subjects with MD-2 promoter SNPs (rs1809441-T/rs1809442-G) than in those with the genotype (rs1809441-G/rs1809442-C) after Der p 2 stimulation, suggesting that subjects with MD-2 promoter SNPs display more effective responses to Der p 2 in the binding activity of GATA-3 (Fig. 6B). The results of flow showed that more increased percentages of IL-4+/CD4+ cells after Der p2 stimulation were observed in subjects (patient Nos. 3 and 4) with MD-2 promoter SNPs (rs1809441-T/rs1809442-G) (+) than in those with SNPs (-) (rs1809441-G/rs1809442-C). The percentage of IL-4+/CD4+ cells in subjects (patients Nos. 3 and 4) were 10.65% and 15.95%, respectively, before Der p 2 stimulation, and then 30.00% and 28.99%, respectively, after the Der p 2 stimulation, suggesting that MD-2 promoter SNPs (rs1809441/rs1809442) may affect the binding affinity of GATA-3 and TH2 cytokine expression.
DISCUSSION
In our previous study, the minor allele frequency of the MD-2 promoter SNPs (rs1809441-T/rs1809442-G) was 0.30 in allergic subjects and 0.17 in healthy subjects. The T allele of rs1809441 and the G allele of rs1809442 were found to be significantly higher in allergic subjects compared to healthy subjects (P<0.05).12 In this study, PBMCs with MD-2 promoter SNPs (rs1809441/rs1809442) were found to have an increased expression of MD-2 and increased secretion of inflammatory cytokines by Der p 2 stimulation. These results indicate that SNPs may be associated with the development of MD-2-related inflammatory diseases. This finding is similar to that of a previous study reporting that SNPs in MD-2 promoter increases MD-2 mRNA production, which correlates with the incidence of sepsis after major trauma.13
Mechanisms underlying the up-regulation of MD-2 expression by Der p 2 cannot be simply due to the structural homology of Der p 2 and MD-2. Der p 2-associated allergic diseases are Th2-dependent, and Th2-cell activation has been shown to be associated with GATA-3 gene expression. Based on the TRANSFAC MATRIX TABLE transcription factor binding site database, sequence fragments of rs1809441 and rs1809442 highly correlate with binding to the transcription site of GATA-3. These results suggest that Der p 2 may trigger MD-2 promoter, followed by activation of GATA-3 and Th2 cells. MD-2 has been reported to play an important role as a cofactor with cell surface TLR-4 and in the recognition of LPS, and group 2 allergenic components of HDM Der p 2 have been reported to serve as an accessory protein in TLR-4 signaling. Therefore, both LPS and Der p 2 may affect host cells synergistically. In our PBMC mRNA expression study, IL-1β and IL-8 were up-regulated by Der p 2 in all subjects, and especially in those with 2 SNP mutants, suggesting that Der p 2 can trigger innate responses in subjects with susceptible MD-2 promoter. The PBMC activation and cytokine (IL-1β and IL-8) release augmented by Der p 2 in these subjects may also have been attributed to the overexpression of MD-2. Similar findings have been reported in which MD-2 expression was up-regulated by Der p 2 in B cells,10 which indicates that the up-regulation of cytokine (IL-1β and IL-8) release may be due to an increased expression of MD-2.
When Cε and Iγ1/2-Cµ mRNA expressions were analyzed, there was no significant augmentation by Der p 2, although the mRNA level of Cε was higher and the level of Iγ1/2-Cµ was lower in subjects with MD-2 promoter SNPs. Therefore, Der p 2-triggered MD-2/TLR4 signaling may only play a partial role in Der p 2-specific IgE synthesis. Other TLR signaling pathways, such as TLR-2 signaling, and other susceptible genes may also be involved in Der p 2-induced B-cell activation and IgE synthesis. According to data from a publicly available database (dbSNP, http://www.ncbi.nlm.nih.gov/SNP), allelic frequencies of genetic variations in the MD-2 or FcεRIα gene promoter region from individuals of different ethnicity are not identical (for example, re1809440 C-allele: 0.479 in African Americans and 0.325 in Europeans). These differences may result in diverse genetic roles of these polymorphisms in different populations with regards to occurrence of certain diseases. Therefore, additional studies are needed to clarify the precise role of these polymorphisms in the pathogenesis of Der p 2 allergy and specific IgE synthesis.
Inconsistent findings on the relationship between exposure to endotoxin and allergic asthma have been reported. It has been hypothesized that LPS can trigger a Th1 response and avoid Th2 prime allergic inflammation. A recent meta-analysis of the association between endotoxin exposure and wheezing/asthma
in children reported a positive association between endotoxin concentration and wheezing in young children, and an inverse association in older children.17
In our cytokine secretion study, PBMCs derived from subjects with MD-2 promoter SNPs (rs1809441/rs1809442) responded differently from those derived from wild type MD-2. Der p 2 could selectively modulate the secretion of the cytokines IL-6, IL-8, and IL-10, and LPS could up-regulate IFN-γ and down-regulate IL-4 secretion, indicating that LPS can prevent Th2-prone allergic responses and that Der p 2 may initiate innate inflammatory responses and contribute to allergic reactions. These findings suggest that both LPS and Der p 2 can trigger inflammatory responses. However, the predetermined genetic background may also play a role in the pathogenesis of allergic inflammation.
It has been reported that the allergenicity of Der p 2 and functional mimicry with MD-2 can facilitate signaling through direct interactions with the TLR4 complex in the absence of MD-2. It has also been reported that allergens from HDMs can induce asthma via TLR4 that triggers airway structural cells to produce innate inflammatory cytokines. This suggests that Der p 2 may activate structural cells directly through its interaction with TLR4 or indirectly through up-regulation of MD-2.1819 In our study, the significantly higher levels of cytokine secretion were found in PBMCs derived from subjects with MD-2 promoter SNPs (rs1809441-T/rs1809442-G) compared to those with the wild type (rs1809441-G/rs1809442-C). Der p 2 could also up-regulate the expression of MD-2. These results suggest that Der p 2 can render PBMCs vulnerable to activation by environmental pathogen endotoxins. It has been reported that the main mite allergen Der p 2 is structurally homologous to MD-2 (also named as LY96 and a molecule required for LPS binding to TLR-4).8 For this reason, Der p 2 induces allergic inflammation in a TLR4-dependent but MD2-independent manner through activation of airway epithelial cells and production of TH2-associated cytokines.8 In addition, the master regulator of transcription factor for TH2-cell differentiation is GATA-3, which promotes TH2 responses through induction of TH2-cytokine production, selective growth of TH2 cells, and inhibition of TH1 cell-specific factors.2021 In this study, the expressions of Cε and GATA-3 in PBMCs derived from allergic subjects were significantly higher in allergic subjects with MD-2 promoter SNPs (rs1809441-T/rs1809442-G) when compared to those with MD-2 wild-type (rs1809441-G/rs1809442-C) both in the conditions of PBMC cultured without and with Der p 2 stimulation. This suggests that MD-2 promoter SNPs may be associated with highly increased expression of the transcription factor GATA-3 and enhanced TH2 cell differentiation.
In conclusion, the results of this study suggest that high secretion levels of proinflammatory cytokines may be predetermined by MD-2 promoter SNPs (rs1809441/rs1809442), which are further up-regulated by Der p 2 and that both genetic and environmental factors can synergistically contribute to allergic inflammation.




 XML Download
XML Download