

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a chronic and complex disorder of the respiratory system characterized by airway obstruction and inflammation,1 and its prevalence is increasing in both developed and developing countries.2,3 This increased prevalence may reflect increased exposure to environmental risk factors. Although environmental factors are important determinants of asthma, numerous studies have revealed that asthma has a strong genetic component. Susceptibility genes have been identified by candidate gene association studies, genome-wide linkage studies, and genome-wide association studies.4 Many genes have been shown to make small contributions to the overall phenotype.
Toll-like receptors are an essential family of innate immune pattern recognition receptors that play a pivotal role in host defense against microbes.5 TLRs can modulate the immune system through cellular activation, modulation of cytokine secretion6 and production of soluble factors to local dendritic networks.7 Human epidemiological studies have supported the common clinical perception that TLRs are associated with asthma. These studies have varied in design, population composition, asthma definition and size. Macrophages, one of the main immune cell types involved in asthma, express various TLRs, including TLR2, 4, 5, 6, 7, 8, and 9.8 Each TLR recognizes different classes of molecules expressed by pathogens and recognizes structurally conserved motifs.9
TLR1 and 6 recognize multiple diacyl peptides, and TLR1/2 dimers can recognize lipopeptides.10 TLR2, which has been found to be expressed by various cell types,11 is related to protection against allergies and allergic asthma by sensing pathogen-associated patterns in lipoproteins and lipopeptides. The TLR2/6 dimer is involved in the recognition of diacylated lipopeptides.12 Long-term stimulation of TLR3 upregulates the production of inflammatory cytokines and cellular recruitment to the airways.13 TLR4 encodes a macrophage cell-surface receptor, which is principally activated by bacterial endotoxin, also known as lipopolysaccharide (LPS). Few studies of the role of TLR5 in asthma have been reported. However, a significant decrease in the expression of TLR5 was observed in asthma patients,14 and TLR5 recognizes bacterial flagellin when complexed with TLR4.15 Similar to TLR5, few data exist on the role of TLR6 in allergic asthma.12 TLR6 is expressed on mast cells, which play important roles in allergy.16 TLR7 and TLR8, which are localized on the sex chromosome Xp22,17 recognize single-stranded RNA, and induce interferons (IFNs) to protect the host from viral infection.18 TLR9 detects bacterial or viral DNA with unmethylated cytosine and guanine.8 TLR10 is the most recently identified gene in the TLR family whose product recognizes pathogen-associated molecular patterns (PAMPs).19 TLRs are also involved in various signaling pathways of the immune system that protect from asthma or develop asthma phenotypes.20,21,22,23,24,25
Most studies published since 2004 have comprised genetic association investigations in various clinical settings, and have found positive associations of SNPs in TLR2,26,27,28 TLR4,29 TLR6,30 TLR7/8,31 TLR9,32 and TLR1019 with asthma. Recent insights into the complex mechanisms of human innate immunity have suggested that genetic variability in genes may play a role in the development of asthma and related diseases.34,35,36,37,38
Due to the important contribution of TLR genes to asthma, an updated meta-analysis was conducted to estimate the effect of SNPs in TLR genes on asthma susceptibility.
MATERIALS AND METHODS
Identification of eligible studies
The review process followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses guidelines.39 We performed a literature search using the MEDLINE and EMBASE databases to identify articles that examined associations between TLR polymorphisms and asthma. Combinations of keywords, such as 'TLR', 'polymorphism', and 'asthma' were entered as medical subject heading (MeSH) and text words. References in the identified studies were used to identify additional studies not indexed by MEDLINE or EMBASE databases. No language or date restrictions were applied.
Inclusion and exclusion criteria
Studies identified from the searches were screened and excluded from further analysis if one of the following reasons was satisfied: a review article, lack of information, animal research, not case-control or nested case-control study design, or unreported genotype frequencies.
For inclusion, the studies must have met the following criteria: (1) they evaluated TLR gene polymorphisms and asthma; (2) were case-control studies; (3) supplied the number of individual genotypes in asthmatic cases and in controls, respectively; or (4) they had an asthma outcome definition that followed accepted diagnostic guidelines.
Data extraction
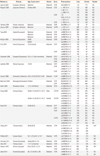
Information was extracted carefully from all of the eligible publications independently by 2 authors, based on the inclusion criteria above. The following information was extracted from each study: author, year of study publication, ethnicity of the study population, demographics (age), asthma status, and number of cases and controls for each TLR polymorphism.
Statistical analysis
Data from the studies were combined to provide a summary odds ratio (OR). Summary ORs were represented as a point estimate with 95% confidence intervals (CIs) on a forest plot.40 The heterogeneity of the data was evaluated using the Q statistic.41 The stability of the summary risk estimate was evaluated using a sensitivity analysis in which each study was removed individually, and the OR was recalculated. Egger's regression test was used to identify publication bias.42
RESULTS
Study inclusion and characteristics
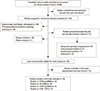
The initial search using the headings 'TLR', 'polymorphism' 'asthma', and their combinations resulted in approximately 6,395 journal articles. Based on the titles, 110 were potentially relevant. Nineteen studies investigating general atopic and allergic phenotypes were excluded. An additional 25 studies on TLR polymorphism involvement in signaling pathways were excluded. Review articles (n=28) were also excluded. Seventeen studies were excluded because they did not meet the inclusion criteria: eight did not report genotypic frequencies in cases and/or in controls24,25,38,43,44,45,46,47; two were cross-sectional studies48,49; four were family-based studies,50,51,52,53 and 3 were abstracts.54,55,56 Twenty case-control studies were eligible for meta-analysis,19,26,27,28,29,30,31,32,33,57,58,59,60,61,62,63,64,65,66,67 one study was in Portuguese,62 and another was in Chinese.64 Among the eligible studies, 11 were excluded because the number of studies of the same polymorphism was less than 3.
The controls for the studies by Lachheb et al.32 and Zhang et al.29 deviated from Hardy-Weinberg equilibrium (HWE) in the controls (P<0.01). The study by Caravalho et al.61 was small, and the definition of asthma was restricted to severe asthma with fungal sensitization. In the study by Palikhe et al.,65 the definition of asthma was limited to aspirin-tolerant asthma. In the study by Yang et al.,58 three sets of patients (first-affected sibling, second-affected sibling and asthmatic parents) were compared with hyper-normal controls. A meta-analysis was conducted when at least 3 studies on the same polymorphism were available. Fig. 1 provides a summary of the search results. Table 1 summarizes the characteristics of the case-control studies.
Data analysis
Association of TLR4 Asp299Gly with asthma risk
Analyses in the random effects model showed that the TLR4 Asp299Gly polymorphism was marginally associated with asthma under the recessive (0.069) (Fig. 2) and allele contrast (0.055) models. In the allele contrast model, sensitivity analysis by exclusion of any one particular study60 in the influence analysis significantly changed the results. The recalculated OR was 0.803 (95% CI, 0.647-0.997; P=0.047), indicating that the major A allele might protect against asthma (Table 2).
The estimated OR1 (AA vs GG), OR2 (AG vs GG) and OR3 (AA vs AG) were 0.897, 0.831, and 0.972, respectively. These estimates were likely to suggest a codominant genetic model. The pooled OR1 was 0.897 (95% CI, 0.597-1.346; P=0.599). Heterogeneity was absent (0.00%). No evidence of publication bias was detected by Egger's test (P=0.12). Sensitivity analysis by the removal of one study did not significantly change the results (Table 2).
Association of TLR4 Thr399Ile C>T with asthma risk
Analyses in the random-effects model showed that the TLR4 Thr399Ile polymorphism was not associated with asthma under any of the genetic models.
The estimated OR1 (CC vs TT), OR2 (CT vs TT) and OR3 (CC vs TC) were 1.223, 1.306, and 0.398, respectively. These estimates did not fit any genetic model. Analysis in the allele contrast model showed no significant association of the TLR4 Thr399Ile C allele with asthma risk (OR=1.223 [95% CI, 0.655-2.286; P=0.528]) (Fig. 3). Heterogeneity was absent in all of the models (0.00%). Sensitivity analysis by the removal of one study did not significantly change the results. Publication bias was detected by Egger's test in the dominant and CC vs TC codominant models (P=0.05 and P=0.03, respectively) (Table 2).
Association of TLR2 Arg753Gln G/A with asthma risk
Analyses showed that the TLR2 Arg753Gln polymorphism was not associated with asthma under the recessive, homozygous (Fig. 4) and allele contrast models. In the homozygous model, sensitivity analysis by the removal of one study61 significantly changed the results. The recalculated OR was 4.460 (95% CI, 1.213-16.396), indicating that the homozygous genotypes might be a significant risk factor (P=0.024). In the GG vs GA model, when one study was removed,61 the recalculated OR was 4.365 (95% CI, 1.227-15.536), suggesting that the homozygous genotype might be a significant risk factor (P=0.023). Sensitivity analysis by the removal of one study60 in the allele contrast model significantly changed the results. The recalculated OR was 0.402 (95% CI, 0.199-0.811), indicating that the major G allele might protect significantly against asthma (P=0.011).
The estimated OR1 (GG vs AA), OR2 (GA vs AA) and OR3 (GG vs GA) were 0.102, 0.01, and 2.451, respectively. These estimates did not fit any genetic model. Only three studies were available; therefore, estimates were not sufficiently powerful. However, these estimates provided a first indication that genotypes with at least one copy of the major G allele might be protective. No evidence of publication bias was detected by Egger's test in all of the genetic models (Table 2).
Association of TLR9 -1237 T/C with asthma risk
Analyses showed that the TLR9 -1237 T/C polymorphism was not associated with asthma under the recessive, homozygous and codominant models. In the allele contrast model, the major T allele was marginally associated with asthma with OR=0.689 (95% CI, 0.471-1.007; P=0.055), suggesting that the major T allele might be a significant protective factor (Table 2 and Fig. 5).
The estimated OR1 (TT vs CC), OR2 (TC vs CC) and OR3 (TT vs TC) were 0.408, 0.01, and 0.498, respectively. These estimates likely suggested a codominant genetic model. Only three studies on TLR9 -1237 T/C were available; therefore, estimates were not sufficiently powerful. However, these estimates indicated that the TT genotype tended to be a significant protective factor (P=0.065). No evidence of publication bias was detected by Egger's test in all of the genetic models (Table 2).
DISCUSSION
During the last decade, the function of TLRs in asthma pathogenesis has been investigated largely by comparing the incidence of disease among persons with different polymorphisms. The magnitude of the association of polymorphisms with asthma varies depending on genetics, demographics, environmental factors, and study methodologies. Many association studies have reported that TLR polymorphisms predispose to asthma.26,47,49,52,56,59,61,68 However, some polymorphisms might not be associated with asthma susceptibility, but rather with asthma severity, such as TLR4 polymorphisms.48,51,58 Conflicting results were also reported, demonstrating a protective effect of some polymorphisms.12,45,46 However, other studies reported no association between TLR polymorphisms and asthma risk.44,50,53,60,61,64,69,70 Given the conflicting results yielded by genetic association studies, a systematic review and meta-analysis is of great value. The current meta-analysis provides a comprehensive examination of the available evidence concerning the association of SNPs in TLR genes with asthma and refines their risk profiles. This analysis represents the first meta-analysis concerning the impact of some TLR polymorphisms on asthma and assesses their role as modifiers of asthma risk.
Estimates of combined effect sizes revealed that the major TLR9 -1237 T allele was significantly associated with a decreased asthma risk. Additionally, for TLR2 Arg753Gln G>A, sensitivity analysis in the allele contrast model revealed that the major G allele was significantly associated with a decreased asthma risk. For TLR4 Asp299Gly A>G, the AA homozygote genotype and A allele tended to be associated with a decreased asthma risk. These results implicate that the major allele in TLR polymorphisms might be generally associated with a protective effect. However, TLR4 Thr399Ile showed no significant association with asthma risk. In a previous meta-analysis, Chen71 reported no direct association between TLR4 Asp299Gly and asthma. This result is consistent with our findings in the dominant, homozygous and codominant models. In a recent systematic review, Klassen et al.72 reported that TLR 2, TLR6, TLR9, and TLR10 appear to have some association with childhood asthma in Caucasians. However, no convincing evidence for a role of TLRs in relation to childhood asthma exists. Polymorphisms in TLR3 seemed not to be associated with asthma.57,65 Results for TLR6 are conflicting. One study with a small sample size showed that TLR6 Ser249Pro C>T contributed to asthma susceptibility,30 but a larger study on the same polymorphism reported no association in both children and adults.59 However, other studies reported that polymorphisms in TLR6 were significantly associated with asthma phenotypes.44,45,56,73 Similarly, some polymorphisms in TLR7/8 have been shown to be associated with asthma disease,29,33 whereas other polymorphisms showed no association.29 For TLR10, some polymorphisms showed significant association with asthma23,33,45; however, other polymorphisms showed no significant association.27,45 Genetic association studies on TLR1 and TLR5 remain scarce or are lacking. Further investigations on TLR polymorphisms, including haplotype analysis along with gene-environment interaction, are needed.
SNPs, which occur at every 1.9 kb in the genome on average,73 could result in reduced or increased gene expression when they occur in promoter.74 This meta-analysis did not control for multiple known risk factors for asthma, such as age, gender, and ethnicity. These potential confounding factors may interact with TLR polymorphisms to moderate asthma susceptibility. Genetic associations were shown to be significantly influenced by age75,76,77,78 likely because some genes are expressed in adults and not expressed in children and vice versa. Observational studies showed that the prevalence of asthma has increased during the last decade due to environmental factors.3 Therefore, it is important to investigate other polymorphisms in TLRs, as well as gene-gene and gene-environment interactions. A possible gene-environment interaction has been reported,79 in which the SNPs act as modifiers of asthma risk in individuals with various degrees of environmental endotoxin exposure. Future studies should include haplotype analysis and stratification of the study population according to related environmental risk factors.
Asthma is a complex disease involving several genes that might be more relevant than TLR genes. Although statistically significant, the effect of the TLRs on asthma was modest. This result may reflect the important role of other confirmed candidate genes in asthma disease. It has been reported that many genes are involved and make small contributions to the overall asthma phenotype.
The findings of our meta-analyses are consistent with experimental studies demonstrating that TLR polymorphisms are important factors in asthma disease, and support the need for further research into the mechanisms underlying asthma susceptibility. The significant association between TLR polymorphisms and asthma risk may be of clinical and public health importance. Further meta-analyses and studies on the expression of related genes, environmental factors, and gene-gene and gene-environment interactions would provide useful information for the prevention and treatment of asthma.
The recent discovery of TLRs, with their role as initiators of the innate immune response and inflammation, suggests that modulating these receptors may be beneficial in the treatment of allergic disorders.7 TLRs are currently being exploited as possible targets for drug development.80 Recently, several patents aimed to modulate the innate immune reaction occurring in asthma through the use of novel synthetic TLR2 ligands8,11; these open new therapeutic perspectives for the prevention of these pathologies. Greater knowledge of risk factors will translate into improved diagnosis, prevention and therapeutic strategies for this chronic disease.





 XML Download
XML Download