

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Eosinophils play an important role in the development of asthma and allergic disease as an effector cell. The extent of eosinophilic inflammation is a determinant of the severity of asthma symptoms,1 and it usually correlates with airflow limitations.2 Relationships between eosinophilic infiltration and Th2 cytokines have been identified within the target tissues of allergic disorders.3 The processes of eosinophilic infiltration and peripheral eosinophilia are dependent on eosinophil-specific cytokines and chemokines (e.g., IL-5; the eotaxin family; RANTES; and MCP-2, -3, and -4), which provoke an eosinophilic response in the peripheral blood and airways via CCR3.4,5 CCR3 mRNA and protein levels are elevated in the bronchial mucosa of asthmatics, and this elevation is associated with airway hyperresponsiveness.6 The participation of CCR3 in airway eosinophilic infiltration has been demonstrated in a study of CCR3-deficient mice, which showed that most eosinophils are arrested in the subendothelial space.7 Eosinophils are derived from CD34(+) hematopoietic progenitor cells.8 Eotaxin-1 and -2 induce the migration of bone marrow and blood CD34(+) CCR3(+) cells in vitro.9 Therefore, the CCR3-eotaxin pathway is important in the regulation of allergen-induced hematopoiesis and in the accumulation and mobilization of eosinophil lineage-committed progenitor cells in the lung.9 The human CCR3 gene (MIM #601268) is located on chromosome 3p21.3.10 The single-nucleotide polymorphisms (SNPs) CCR3-22557G>A and -174C>T are associated with the number of eosinophils in asthmatic patients. It has previously been reported that CCR3 expression is higher on the eosinophils of asthmatic patients lacking a haplotype composed of rare alleles of 4 CCR3 SNPs.11
Eotaxin-1 is important for eosinophilic inflammation in early stages of the asthmatic response, while eotaxin-3 may account for eosinophil recruitment to the airways in late stages of the asthmatic response.12 Three eotaxin family members, eotaxin-1, -2, and -3, are selectively bound by CCR3. Eotaxin-1 and -2 show different pathophysiologic responses in relation to eosinophils. Disruption of the eotaxin-1 gene leads to a slight reduction in the eosinophil count in the blood and airways.13 Eotaxin-2-deficient mice have normal baseline eosinophil levels in their target tissues, and they do not develop airway eosinophilia in response to IL-13.14 Two eotaxin polymorphisms, EOT2+1272A>G and EOT1+123G>A, are associated with asthma and high serum total IgE levels, respectively.15
Because asthma is a multifactorial disease, the genetic component may be derived from the combined effect of numerous genes. Individual genes may act independently or in combination with other genes in the same biological pathway, resulting in variable effects.16 The CCR3 gene and eotaxin gene family are possible contributors to the development of asthma or other allergic phenotypes via the receptor-ligand interaction.
In this study, we evaluated gene-gene interactions between CCR3 and eotaxin in relation to eosinophilia in patients with asthma.
MATERIALS AND METHODS
Subjects
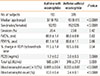
We enrolled 533 Korean asthmatics in our study. The Institutional Review Board of [Soonchunhyang University Bucheon Hospital] approved the study. All of the patients had current symptoms, including wheezing, dyspnea, and cough, and met the criteria for asthma as defined by the American Thoracic Society.17 Each patient showed airway reversibility as documented by an inhalant bronchodilator-induced improvement in the forced expiratory volume in one second (FEV1) of >15%18 and/or airway hyperresponsiveness as shown by a provocative concentration of methacholine required to cause a 20% decrease in the FEV1 (PC20) of <8 mg/mL.18 The asthma subjects were divided into 2 groups according to the presence of eosinophilia. Eosinophilia was defined as a blood eosinophil count exceeding 0.5×109/L.19 No subject had used systemic or inhaled steroids for 4 weeks before peripheral blood eosinophil counting. No subject had a history of medication or the ingestion of raw foods that could produce blood eosinophilia. ELISAs for 4 common parasites (Paragonimus, Clonorchis, Cysticercus, and Sparganum) were negative in the enrolled patients. The clinical characteristics of the subjects are summarized in Table 1.
Genotyping by single-base extension and electrophoresis
To genotype polymorphic sites, amplifying and extension primers were designed for single-base extension. All primer extension reactions were performed using a SNaPshot ddNTP Primer Extension Kit (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions. Detailed methods are described in the online supplement.
Analysis of polymorphisms, Hardy-Weinberg equilibrium, and linkage disequilibrium
A total of 14 SNPs in eotaxin-1, -2, and -3 (6 SNPs in eotaxin-1, 5 in eotaxin-2, and 3 in eotaxin-3), and 4 SNPs in CCR3 were included in our analysis. Testing for Hardy-Weinberg equilibrium and the calculation of D' for the identification of linkage disequilibrium were performed using PHASE v2.0.220 and Arelquin v2.0.21 Continuous variables, such as the blood eosinophil count and percentage, serum total IgE level, and PC20 methacholine level, were log-transformed to approximate normal distributions. Differences in the log-transformed values among genotypes were examined using a generalized linear model type III SS. Statistical significance was defined at the standard 5% level.
Dependency of allele frequency and logistic regression interaction modeling
The dependency between each pair of CCR3 and eotaxin SNPs in asthmatics with and without eosinophilia was tested by Chi-square analysis.
An epistatic model was designed based on the logistic regression model.22 "CCR3eff," a new numerical variable as an effect of CCR3 SNPs on blood eosinophilia, was derived according to the extent of contribution to eosinophilia in asthma using logistic regression. Odds ratios (ORs) for eosinophilia for each genotype at each locus were calculated in asthmatics with eosinophilia. Each genotype was coded according to the ORs as follows: homozygote with the highest OR, 2; heterozygote, 1; and the other homozygote, 0.
In CCR3eff=α1 (-22557G>A)+α2 (-520T>G)+α3 (-174C>T)+α4 (+51T>C),
α denotes the coefficient fitted by logistic regression (α1=0.3601, α2=0.3525, α3=0.4204, and α4=0.1867). Epistasis was subsequently tested independently for each SNP in eotaxin by fitting the following logistic regression model:
P (Eosinophilia)=exp (X)/[1+exp (X)]
where X=β0+β1CCR3eff+β2SNPEotaxin+β3 (CCR3eff×SNPEotaxin), β0 is the intercept, β represents the parameter estimates, and SNPEotaxin is each eotaxin genotype (0, 1, or 2).
The Bonferroni correction for multiple comparisons was used for each eotaxin gene (the global significance level was adjusted to P<0.0083 for 6 eotaxin-1 SNPs, P<0.01 for 5 eotaxin-2 SNPs, and P<0.017 for 3 eotaxin-3 SNPs). Statistical significance was defined at the standard 5% level.
Generalized multifactor dimensionality reduction (GMDR)
To validate the results of our logistic model, we performed GMDR analysis. First, the data were randomly split into 10 equal parts for cross-validation. Nine of these were used as a training set; the remaining set was used for independent testing. Cross-validation is a measure of the number of times a particular set of loci is identified in each possible 9/10 of the subjects. Second, a set of n genetic factors was selected. Third, all possible multifactor classes or cells were represented in n-dimensional space and a cumulative score was calculated within each cell.23 Fourth, each multifactor cell in n-dimensional space was labeled as "high-risk" if the average score met or exceeded the threshold (0) or as "low-risk" if the threshold was not exceeded. This process was repeated for each possible cross-validation interval. Fifth, all potential combinations of n factors were evaluated sequentially for their ability to classify cases and controls in the training data, and the best n-factor model that yielded the minimum misclassification error was chosen. Sixth, the independent testing set was used to estimate the prediction error of the best model selected in the fifth step. Finally, among this set of the best models, we pick the model with minimum prediction error and/or maximum cross-validation consistency.23,24 A covariate analysis was performed as described above23 with age (continuous variable) and sex (discrete variable) as covariates.
RESULTS
The genotype distributions of the four SNPs in CCR3 and 14 SNPs in the eotaxin genes were in the Hardy-Weinberg equilibrium (data not shown). No pair of SNPs showed strong linkage disequilibrium (>0.8) between the eotaxin gene SNPs and CCR3 SNPS (see Supplementary Table S2).
Epistasis was evaluated to access whether the eotaxin SNPs modified the effect of the CCR3 SNPs on blood eosinophilia susceptibility. We examined whether dependence existed between the CCR3 SNPs and eotaxin gene SNPs before the epistasis tests. EOT2+304C>A was dependent on 3 intronic SNPs (CCR3-22557G>A, CCR3-520T>G, and CCR3-174C>T) in CCR3 among asthmatics with eosinophilia by Chi-square analysis (Table 2).
The epistatic model constructed by logistic regression that tested the effect of the combined CCR3 SNPs on blood eosinophilia differed depending on the eotaxin SNP: EOT2+304C>A (29L>I) with the CCR3 SNPs was significantly associated with blood eosinophilia (P=0.0087; Table 3). Thus, genetic susceptibility to blood eosinophilia in asthmatics is conferred by both CCR3 and eotaxin.
According to our GMDR results, the combination of CCR3-174C>T and EOT2+304C>A was the best model (accuracy=0.536, CVC 9/10; Table 4). This combination was also the best model after adjustment for age and sex as covariates. The combination of the CC genotype of CCR3-174C>T and AA or AC genotype of EOT2+304C>A was detected in the high-risk group. The combination of CT of CCR3-174C>T and CC of EOT2+304C>A was also detected in the high-risk group (Figure).
DISCUSSION
Given the wealth of data showing that CCR3 and eotaxin participate in eosinophilic infiltration, the cooperative effect of the SNPs of CCR3 and eotaxin on eosinophilia in asthmatics was analyzed in this study. Three SNPs (CCR3-22557G>A, CCR3-520T>G, and CCR3-174C>T) in CCR3 were significantly associated with the number of eosinophils in patients with asthma.11 Associations between EOT2+1272A>G and the risk of asthma, and between EOT1+123G>A and high serum total IgE levels, were reported in Korean asthmatics.15 However, these SNPs were not found as a best single-locus model in relation to blood eosinophilia in this study.
Gene-gene interactions in various diseases have been studied using a variety of approaches. These studies demonstrated that the overall disease risk can be modeled as the product of the risk conferred by many independent risk factors.16 In this study, gene-gene interactions were examined using 3 methods. First, a Chi-square test was conducted. Using this approach, EOT2+304C>A was found to be significantly associated with the 3 intronic SNPs of CCR3 in patients with eosinophilia (Table 2). Next, we used logistic regression analysis and designed an epistatic model as described previously.22 Three of the 4 CCR3 SNPs contributed independently to the eosinophilic phenotype in asthma. This model can be used to evaluate the mean effect of all SNPs in CCR3. Of the 14 SNPs in the eotaxin gene family, only 1 in eotaxin-2 (EOT2+304C>A [29L>I]) was significantly associated with blood eosinophilia; however, the association was modified by the effect of CCR3 SNPs (P=0.0087; Table 3). Finally, we constructed an epistasis model using GMDR with and without covariate adjustment. Because the number of subjects in the 2 groups (asthmatics with or without eosinophilia) was different, GMDR was used instead of regular MDR. The combination of EOT2+304C>A and CCR3-174C>T was the best model. This model was still the best after adjustment for age and sex as covariates (Table 4). We successfully identified genotype combinations that contribute to eosinophilia in asthma using this model. Asthmatics, who had the C genotype of CCR3-174C>T and A or AC of EOT2+304C>A and who had the CT genotype of CCR3-174C>T and C of EOT2+304C>A (Figure), were included in the high-risk group. Thus, the number of blood eosinophils is higher in asthmatics bearing these genotype combinations.
No single SNP in the eotaxin gene family was associated with blood eosinophilia in asthma (data not shown). However, EOT2+304C>A in which leucine is changed to isoleucine interacted significantly with the effect of CCR3 SNPs on blood eosinophilia in asthmatics. This finding remained significant after correction with the Bonferroni test (P<0.01). The eotaxin and CCR3 pathways are thought to interact biologically, suggesting a gene-gene interaction, or epistasis. The term "epistasis" has been used to describe a "non-independence of effect". In essence, if the effect of one unit is not predictable unless the value of another unit is known, the effect is epistatic.25 In this regard, the effect of EOT2+304C>A may not be disclosed if the effect of CCR3 SNPs is not considered. Moreover, there is no definite evidence that gene-gene interactions can be identified only among loci that show a significant association with phenotype.
There are some limitations to this study. First, blood eosinophilia was included in our models instead of sputum eosinophilia; however, a small number of patients had sputum eosinophilia. Second, it is difficult to confirm the results of interaction models biologically. Eosinophils are further decreased in the lungs and peribronchial tissues of eotaxin-1/-2 double-knockout mice, compared with CCR3-deficient mice.26 However, the knockout of eotaxin and CCR3 in animals has not been reported.
In summary, novel gene-gene interactions between SNPs of CCR3 and an SNP of eotaxin-2 in the same phenotype were identified in this study. Our analysis of the epistatic influence of CCR3 and eotaxin gene variants suggests that these variants may be candidate markers for eosinophilia in asthma and could be important in understanding the genetics underlying allergic diseases.




 XML Download
XML Download