

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Epidemiological research suggests that exposure to environmental toxins (such as pesticides, solvents, and air pollutants) is associated with an increased prevalence of asthma and allergies. Asthma and allergic diseases are the most common chronic inflammatory diseases in children, and represent a substantial morbidity and mortality burden in severe cases.1 The prevalence of allergic disorders has seen a worldwide increase over the last few decades. Most countries report population rates of allergic diseases (such as allergic rhinitis and asthma) between 15%-35%, with the worldwide average being 22%.2 Asthma prevalence has been increasing since the early 1980s and by 2004, 1 in 15 Americans suffered from asthma.3 Exposure to environmental toxicants not only contributes to the increasing prevalence of asthma and allergies, it also affects disease outcomes, many of which are due to an underlying immune and inflammatory dysfunction.4 Mounting epidemiological evidence suggests that a proper functioning of the immune system is a major determinant of health across the course of a lifetime; for example, adults who suffered with childhood asthma may have an elevated risk of developing lung cancer.4 Therefore, exposure to environmental toxicants may cause life-long changes in responses to infectious agents and in the homeostasis of the immune system, as well as in our overall physical health. Epidemiological studies have suggested that following exposure to air pollution, variant genes controlling the innate immune response and the subsequent inflammatory reactions, are associated with an adverse respiratory outcome.5 Some environmental toxins, such as endocrine disrupting chemicals (EDCs), imitate the action of steroid hormones and have promoted endocrine and reproductive disorders, both in animal and human studies. These environmental toxins interfere with the synthesis of cytokines, immunoglobulins, and inflammatory mediators, and they also affect the activation and survival of immune cells. The dysfunction of the immune system caused by some EDCs may lead to attenuation of immunity immunodeficiency against infection, or to hyperreactivity of immune responses (allergy and autoimmune disease). In vitro and in vivo studies have demonstrated that many of the environmental chemicals and pollutants that have been associated with increased allergic tendency promote type 2 helper T cell (Th2) reactions, consistent with Th2 predominant responses found in asthma and allergic rhinitis. In this review, we have summarized epidemiologic, animal, and cell studies to demonstrate the impact of indoor and outdoor environmental toxins on allergy diseases and the associated immune mechanisms, such as the regulatory effects of epigenetics.
The effects of toxins found in the indoor environment on allergies
EDCs
EDCs have increased since World War II6, and this coincides with an increased prevalence of autoimmune and allergic diseases.7 EDCs are ubiquitous in the environment, and are found in the air, water, and the soil, and many have undesirable effects on human health. EDCs are regarded as inducers of the inflammatory response, and this can be via the estrogen receptor, or other receptors, such as the aryl hydrocarbon receptor.8 EDCs have also been reported as potential modulators of the immune system and allergic responses in allergic disease.9 For example, a high concentration of diethyl hexyl phthalate (DEHP) in indoor dust is associated with wheezing in preschool children. Another alkyphenol, p-octylphenol, has been shown to enhance strong Th2 polarization via suppression of type 1 helper T cell (Th1) and augmentation of Th2 immune responses, respectively.6,10 Recently, new evidence has shown that exposure to alkylphenols plays a key role in the allergic response that may be associated with the development of asthma. Due to the low solubility, high hydrophobicity, and low estrogenic activity, alkylphenols tend to accumulate in the human body and lead to promote the development as well as progression of allergic diseases. Alkylphenols exert their effects on several key cell types in the context of allergic inflammation.
Nonylphenol (NP), one of the alkylphenols, is the most important metabolite of a group of nonionic surfactants, designated as NP polyethoxylates11 and is one of the common EDCs.12 NP decomposes in the environment, and it can influence human health via bioaccumulation in the diet or through other contact means.13 NP is structurally similar to 17β-estradiol, which feminizes male animals and has possible links to infertility.14 Some in vitro or ex vivo studies suggest that NP skews T cells towards Th2 responses through its influence on dendritic cells (DCs). For example, splenic conventional DCs from NP-exposed mice have shown a potent Th2-skewing ability and express increased levels of IL-6 and TNF-α, but not IL-10 and IL-12, in response to LPS stimulation.15 Further, bone marrow-derived DCs in the presence of NP can influence antigen-specific T cells to secrete significantly less IFN-γ.15 Importantly, NP-exposed mice developed relatively more severe OVA-induced allergic lung inflammation.15 Plasmacytoid DCs (pDCs) are the predominant cells that secrete type I IFN during infection. Type I IFN increases the survival of T cells, the differentiation of Th1 and cytotoxic T lymphocytes, and also increases the activity of natural killer cells, leading to an overall boost in the antiviral T-cell activity.16 Concurrently, type 1 IFNs suppress the Th2 immune response.17 We previously reported that NP and 4-octylpnehol (4-OP) influenced the function of pDCs in vitro and in vivo by inducing the pro-inflammatory cytokine TNF-α, and by suppressing regulatory cytokines such as IL-10, IFN-α and IFN-β, suggesting the potential impact of endocrine disrupting chemicals on immune regulation.18 Accordingly, exposure to NP may be linked to an impairment in anti-viral immunity.
The tendency to develop asthma and other chronic allergic inflammatory disorders such as allergic rhinitis, allergic conjunctivitis and atopic dermatitis, is inherited, but it can be influenced by the environment and modified by in utero exposure and aging. These features are all linked to epigenetic regulation. Environmental exposure, especially prenatal maternal smoking, has been associated with asthma and can be partly explained by epigenetic regulation. There are several forms of epigenetic regulation, including DNA methylation or histone modification. NP increased the expression of TNF-α, but suppressed IL-10 production in a range of physiological doses, concomitant with activation of the MKK3/6-p38 signaling pathway and enhanced levels of acetylated histone 3, and histone 4 at the TNFA gene locus. NP or 4-OP-induced increased TNF-α levels in myeloid DCs (mDCs), which was concomitant with increasing levels of methyltranferase complex mixed-lineage leukemia and tryptophan-aspartic acid repeat domain 5 (WDR5) in the nucleus, that subsequently contributed to the increasing levels of trimethylated H3K4 at the TNFA gene locus. The up-regulated TNF-α expression by NP could be suppressed by a histone acetyltransferase inhibitor. In the presence of NP-treated mDCs, T cells showed increased levels of IL-13 but decreased expression of IFN-γ.19 In a cell study, NP suppressed type I IFN production in response to CpG stimulation, along with down-regulation of IRF-7 and involvement of the MKK1/2-ERK-Elk-1 pathways, leading to the impairment of anti-enterovirus 71 activities. Furthermore, splenic pDCs from NP-exposed mice showed similar cytokine changes upon CpG stimulation under conditions relevant to routing levels of exposure in humans. The exposure to NP promoted allergic lung inflammation in a murine model of asthma.15,18
Phthalates are compounds added to plastics to make them more flexible. Phthalates can easily be released into the surrounding environment because they are not strongly bound to the plastics. At least 2 types of phthalates are reported to be linked with higher rates of allergic respiratory incidences. In a cohort study of 10,852 children, exposure to butyl benzyl phthalate (BBzP) and DEHP was associated with rhinitis, eczema and asthma.20 DEHP is found in all PVC products, and exposure to PVC flooring during pregnancy could be a critical factor in the development of asthma in children.21 Phthalates are reported to enhance Th2 differentiation, the production of Th2 cytokines, and increase levels of Th2-promoted immunoglobulins (mainly IgG1 but also IgE) in mice.22 Similar to NP, phthalates may also interfere with immunity against viral infection and promote a decrease in the Th2 response, thus increasing the allergic reaction by acting on human pDCs via suppression of IFN-α/IFN-β expression and modulation of the ability to stimulate T-cell responses. DEHP and BBP can suppress CpG-induced IFN-α/IFN-β expression in pDCs by suppressing CpG-induced interferon regulatory factor (IRF)-7 expression, via suppression of the histone H3K4 trimethylation at the IRF7 gene promoter region and inhibiting translocation of H3K4-specific trimethyltransferase WDR5 from the cytoplasm into the nucleus. BBP- or DEHP-treated pDCs also suppress IFN-γ while enhancing IL-13 production by CD4+ T cells.23
Tobacco smoke
Cigarette smoke contains harmful chemicals with hazardous effects on almost every organ in the body of smokers, and non-smokers exposed to environmental tobacco smoke (ETS). At least 40,000,000 children are regularly exposed to ETS every year, increasing their risk for acute respiratory infections. It has been recognized that asthma and wheezing are more common in children exposed to cigarette smoke. Tobacco smoke is thought to be the main indoor air pollutant and can lead to higher rates of allergy and asthma, with early exposure to ETS being associated with wheezing in children.24 Diseases of the airways in later life can often be influenced by environmental exposure that occurred during critical developmental periods of the immune systems.25 Maternal smoking is of greater significance than paternal smoking and prenatal exposure is also more significant than postnatal exposure for childhood allergic sensitization.26 Exposure to maternal smoking prenatally and after birth also increased the risk of wheezing and asthma in children.26 Thus, avoiding tobacco smoke exposure during pregnancy and in early childhood might prevent or delay the development of asthma.27 Passive smoking also increases the incidences of wheezing and asthma in children and young people by at least 20%. Therefore, stopping parents smoking is crucially important to the prevention of childhood asthma.28 Second-hand smoke exposure has a dose-dependent synergy with a family history of asthma. Avoiding second-hand smoke may be an important preventative method for reducing the risk of adult-onset asthma among those with asthmatic heredity.29 Detectable levels of serum and salivary cotinine, a marker for tobacco smoke exposure, are common in children admitted for asthma.30 Detectable serum and salivary cotinine levels are also associated with re-admission in children hospitalized for asthma.30 There is evidence to support an association between the development of asthma and tobacco smoke exposure, and evidence is mounting to suggest that exposure to tobacco smoke may also increase the risk of IgE sensitization.31 Tobacco smoke reduces the Th1 response by suppressing IFN-γ production and natural killer cell activity, but it enhances the Th2 response by increasing production of IL-4, IL-5 and other pro-inflammatory cytokines that result in increased allergic responses.32,33,34
Heavy metals
There is an association between heavy metals and allergic reactions. Prenatal and early postnatal exposure to heavy metals, such as lead and mercury, has been reported to skew immune responses toward a Th2 bias and elevate production of IgE or Th2-related cytokines. Lead exposure in mice promotes a Th2 reaction and suppresses IFN-γ expression.35 Mercuric chloride induces a higher IgE production and increases IL-4 levels.36 In a study of children, increased ambient nickel and vanadium concentrations were significantly associated with an increased probability of wheezing at 24 months old.37
The effects of toxins found in the outside environment on allergies
Diesel exhaust particles
Epidemiologic studies have suggested that prenatal and early childhood exposure to diesel exhaust plays a role in the development of allergies. The offspring of mice exposed to diesel exhaust particles (DEP) were hypersensitive to ovalbumin (OVA), indicated by inflammation of the airways and hyperresponsiveness, increased serum OVA-specific IgE levels, and increased pulmonary and systemic Th2 and type 17 helper T cell (Th17) cytokine levels.38 However, exposure to DEPs alone did not induce asthma in mice. The co-exposure of DEPs and house dust mites (HDM) markedly enhanced airway hyperresponsiveness compared with HDM exposure alone and induced Th2 and Th17 responses, including IL-13+, IL-17A+ double-producing T cells.39 In a recent study of children with allergic asthma, those exposed to high levels of DEPs had more frequent symptoms over a 12-month period (32.2%) compared with only 14.2% in a low DEP-exposure group. Children with allergic asthma exposed to high DEP levels had ~6 fold higher serum IL-17A levels compared with low DEP-exposed children.39
Particulate matter
Particulate matter (PM) is a heterogeneous mixture of small particles and liquid droplets suspended in the air. PM can be classified into three categories based on the particle size. Coarse PM (diameter, 2.5-10 µm) is derived from abraded soil, road dust, construction debris and an aggregation of smaller combustion particles. Fine (≤2.5 µm) and ultrafine (≤0.1 µm) PM is formed during the combustion of fossil fuel products. Diesel exhaust is a major source of particulates in urban areas, and accounts for most airborne PM. Particulates can stimulate our immune system to induce inflammatory reactions. Several nanometer-to micrometer-sized particulates, such as particle matter 2.5 (PM2.5), diesel particles, and sand dust, induce pulmonary inflammation, and especially allergic asthma. Exposure to PM air pollution induces an exacerbation of asthma in children, and their exposure to PM is likely to include combustion-related ambient PM and PM related to the child's activity in their indoor and outdoor microenvironments. PM can pass through the throat and nose and enter the lungs, and has an adverse effect on many organs in the body. PM size may be directly linked to its potential to induce immune responses and cause allergy problems. PM2.5 may be more respirable and hence more toxic. There is an association between ambient PM2.5 and the development of allergic asthma and exacerbation of acute asthma. There is also increasing evidence to link ambient levels of PM2.5 to declining lung function, and increased reliance on the need for medical services to treat asthma.40,41,42 PM components from fossil fuel combustion can induce oxidative stress initiated by reactive oxygen species (ROS). Reported associations between worsening asthma and PM2.5 mass could be related to the oxidative potential of PM to induce airway inflammation as a result of oxidative stress. DEPs contribute significantly to urban PM2.5 concentrations.43 The decrease in the FEV1 of asthma patients was significantly associated with increased hourly peaks and the daily average levels of PM2.5.40,44 Nanometer- to micrometer-sized crystals, spheres, and hydrogel forms of aluminum salts have been used as vaccine adjuvants to enhance antibody responses, and most of these particulates induce Th2 immune responses.45
Several particulates exhibit adjuvant effects in immune responses For example, aluminum can selectively stimulate humoral immune responses, especially Th2 immune responses, which are characterized by the production of IL-4 and IL-5 and the induction of IgE and IgG1.46,47 Crystalline silica can also induce Th2 responses and antigen-specific IgE and IgG1.48 Synthesized particles, such as polystyrene particles, poly lactic-co-glycolic acid, nickel oxide nanoparticles and carbon nanotubes, induce humoral immunity, especially antigen-specific production of IgG1 and IgE.49,50,51 Epidemiologic studies have consistently shown that PM can cause asthma and allergies in children.52,53,54
Pesticides
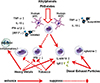
Pesticides are widely used to control pest and pest-related diseases in agriculture, fishery, forestry, and also in the food industry. There are a number of respiratory diseases and symptoms that have been associated with occupational exposure to pesticides. Impaired lung function has also been found in people suffering from allergies who are occupationally exposed to pesticides. Several respiratory symptoms, such as coughing, wheezing and airway inflammation, are commonly observed in people exposed to pesticides.55,56 Exposure to pesticides in utero has been reported as a risk for childhood asthma.57 There is strong evidence of an association between occupational exposure to pesticides and asthma, especially in agriculture. If the fetus is exposed to certain pesticides during pregnancy, allergies and hay fever appear to be more common in the offspring, and especially in the male offspring.57 The potential effects of early exposure to pesticides may be confounded by the generally protective effects of in utero exposure to the farmyard environment and exposure to livestock. Livestock farmers on farms had significantly lower incidences of allergic rhinitis than crop farmers.58 There is little antigenicity in chemical pesticides, pesticide-induced or -promoted allergic/atopic asthma. Therefore, pesticide-induced allergy may be due to the indirect effects of pesticides on the immune system, such as interfering with the Th1/Th2 balance, or pesticide-induced oxidative stress.59 There is an urgent need for training and education on basic safety precautions and the proper use of personal protection equipment. This type of intervention could control the incidences of respiratory disease associated with pesticide exposure in an occupational setting. We summary the potential immunological mechanisms induced by environmental toxicant on allergic inflammation in Table and Figure.
CONCLUSION
Allergic diseases affect a significant percentage of children, and represent the largest group of pediatric chronic diseases in most countries. A wide range of environmental factors and toxins are associated with childhood allergic diseases. Air pollution and toxins should be targeted for intervention as they are a major cause of allergic diseases, particularly in children. Because most of the risk factors are controllable, disease prevention by avoidance of prenatal and neonatal environmental risk factors may offer significant promise to reduce the prevalence of pediatric allergies. However, these established risk factors cannot account for all the recent increases in immune-dependent pediatric asthma. Therefore, new chemical and drug safety testing strategies are needed to identify the main environmental factors and to ensure adequate protection of the most vulnerable i.e., pregnant women and children, from allergy-related developmental immunotoxicity. Early-life immune dysfunction/imbalance has been recognized as a biomarker for the risk of pediatric allergy and asthma. Oxidative stress and epigenetic mechanisms are also considered as possible factors in the development of allergic disease and asthma. Further studies to identify the major environmental risks and to understand how these mechanisms regulate gene-environment interactions will lead us to develop new methods of controlling or preventing allergic disease in children.

 XML Download
XML Download