

 PDF
PDF ePub
ePub Citation
Citation Print
Print


THE SWEET ENEMY IN THE MODERN HOME
"the modern cultivated person sleeps in beds which are a mockery of all the rules of hygiene, ... daily use through dust, fungal spores, bacteria, and from the effect of the damp bed warmth, an opulent breeding ground of fungi, yeasts, and harmful vermin, in beds... All the worse that for sure asthma is not the only illness to arise from mites..."
Hermann Dekker, 1928.1
As early as 1921 it was reported that many asthmatics had strongly positive skin tests to house dust.2 The first attempt to purify allergens from house dust was unsuccessful.3 In 1928, Dr. Dekker suggested that mites played an important role in allergy, but his evidence was unconvincing.1 A breakthrough came in the mid-1960s, when Spieksma and Voorhorst established that there was a strong relationship between the number of dust mites and the allergenicity of house dust.4 Using skin test reagents, it became obvious that there was a strong association between dust mites and asthma. In some areas, the association was so strong that 80%-90% of children with asthma had sensitivity to dust mite allergens.5 In fact, early sensitization to perennial indoor allergens has been implicated in several recent reports as a major risk factor for a persistent type of respiratory allergic disease in childhood.6,7
THE GLOBAL ENDEMIC OF HDM ALLERGY
There is increasing evidence that exposure to indoor allergens is a causative factor for the development of asthma among persons who are genetically predisposed to mount IgE autoantibody responses.8-10 Hypersensitivity to house dust mite (HDM) allergens is one of the most common allergic responses. Sensitization to indoor allergens is the strongest independent risk factor associated with asthma (odds ratios>10).11 In fact, more than 90% of allergic asthmatic children are sensitive to Dermatophagoides spp (D. pteronyssinus, Derp and D. farina, Der f).12 Reports from others also suggest that >50% of children and adolescents with asthma are sensitized to HDM.11,13
GEOGRAPHY OF HDM
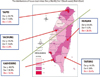
Multicenter studies suggest that HDM are the most prevalent source of indoor allergens in Europe,14 USA,15 Asia,16,17 South America,18 New Zealand,19 Australia,20 and Africa.21,22 The distribution of house dust mites and their species vary around the world according to temperature and humidity.23 A recent survey in Taiwan revealed that, along with D. pteronyssinus (Der p) and D. farina (Der f), the most common mite species in house dust fauna was Blomia tropicalis (Blo t).24 As shown in Fig. 1, there is a significant difference in the distribution of Blo t in house dust between the northwest (3.9%-7.3%), which is cold and humid, and the southeast (18.9%-28.9%), which is hot and humid. Due to the high humidity, the majority of Dermatophagoides species were Der p. The prevalence of Der p in the north and south parts of Taiwan was not markedly different.
THE ALLERGIC IMMUNE RESPONSE TO HDM
Allergens derived from HDM have been recognized as an important cause of IgE antibody responses for more than 30 years. During allergen sensitization and provocation in allergen-induced inflammation, mite allergens break the anatomical barrier of the mucosal membrane and are processed by professional antigen presenting cells (APCs), such as dendritic cells or macrophages. APCs mature and present processed allergen peptides to resting naïve T cells in the draining lymph nodes. Activated T cells signal to APCs, particularly DCs, to produce Th2 cytokines, such as IL-3, IL-4. IL-5, and IL-13, which recruit, mature, and activate eosinophils. These cytokines induce isotype switching from IgG to IgE, leading to increased production of allergen-specific IgE that eventually binds to high affinity IgE receptors (FCεR1) in mast cells, basophils, and eosinophils. Therefore, subsequent encounters with the same allergen will result in degranulation of preformed granules from IgE-IgE receptor-complex-bound mast cells and basophils (immediate hypersensitivity phase), production of newly synthesized inflammatory mediators (late inflammatory phase), and a hypersensitivity response (chronic phase). The innate immune response may also play a role by being induced by the allergen itself.25
THE INNATE IMMUNE RESPONSE TO HDM
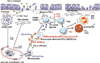
More than 20 HDM allergen groups that induce IgE antibodies in allergic patients have been defined based on sequence and functional homologies.26 When these allergens contact the mucosal membrane, they may induce innate immune stimulation. We have demonstrated that extracts from dust mites (Der f) can directly activate innate immune cells such as alveolar macrophages (AMs),27 and mast cells,28 and can induce Th2 cytokine responses without previous in vitro or in vivo sensitization. Proteases in HDM may induce the innate immune response by binding protease-activation receptors (PARs), Toll-like receptors (TLRs), or C-type lectin receptors (CTRs), or they may function as molecular mimics (Fig. 2), as discussed below.
THE PROTEASE ACTIVITY AND PAR ACTIVATION IN HDM ALLERGY
HDMs and their fecal pellets contain several proteolytic enzymes. Group 1 allergens are cysteine proteases that share sequence identities with the catalytic sites of the plant enzyme papain. Groups 3, 6, and 9 are serine proteases,29,30 which account for 79% of the proteolytic activity in house dust.31 Proteolytically active allergens can down-regulate the anti-protease-based lung defenses in mucosa, leading to enhanced tissue damage and immune activation. Der p 1 and Der f 1 can degrade airway α1-antitrypsin inhibitor, resulting in dysregulation of human neutrophil chemotaxis.32 Previously, we found that pulmonary surfactant proteins A (SP-A) and D (SP-D) secreted from alveolar type II cells, which belong to the innate immune defense in the lung, are able to bind to Der p extracts33 and prevent Der p allergen-induced histamine released from mast cells in sensitized patients.34 These two mite cysteine proteases can inactivate SP-A and SP-D (collectins), which have significant functions in the innate defense mechanism against pathogens. Collectins also facilitate the recognition of aero-allergens, and protect against and resolve allergen-induced airway inflammation.35,36
Der p 1 triggers the breakdown of the epithelial barrier through proteolytic cleavage of tight junction proteins occludin and zonula occludens-1 (ZO-1).37,38 This increase in the permeability of the bronchial epithelium can facilitate allergen presentation by airway DCs in subepithelial tissues.38 Der p 1 downregulates IDO expression in DCs from HDM-sensitive subjects with asthma, which explains the loss of tolerance to HDM allergens.39 Proteolytically active Der p1 also induce airway epithelial cells, skin keratinocytes, DCs, eosinophils, basophils, and macrophages to secrete large amounts of proinflammatory, pro-Th2 cytokines, such as TSLP and IL-33, as well as chemokines to recruit inflammatory cells to the damaged epithelium.40
Group 3 (Der p 3) and 9 (Der p 9) mite allergens are serine proteases that can increase vascular permeability and detach epithelial cells.30 Although the receptors for these proteases have not yet been characterized, it is probable that the primary response is due to activation of cell surface protease-activated receptors (PARs) or similar molecules in the airways. This would induce leukocyte infiltration and amplify the response to allergens.41
PARs are receptors activated by proteolytic cleavage of their extracellular N-terminus. Signaling through PARs (G protein-coupled receptors) involves the cleavage of an extracellular region of the receptor by proteases to reveal a tethered-ligand sequence capable of auto-activating the receptor.42 Protease inhibitors already in clinical use for the treatment of endogenous protease release-related diseases, such as acute pancreatitis, may have some beneficial effect in the inhibition of proteolytically active allergen-induced airway inflammation. In fact, we have found that gabexate mesylate (FOY) and nafamostat mesylate (6-amidino-2-naphthyl p-guanidinobenzoate dimethane sulfonate [FUT]), which are non-antigenic synthetic inhibitors of trypsin-like serine proteases that have been used in the treatment of pancreatitis and disseminated intravascular coagulation, are able to alleviate mite allergen-induced airway hyperresponsiveness and inflammation in the allergic asthma mouse model.43 Moreover, a monoclonal blocking antibody (Wan 108) against Der p 1 developed by our group can not only interfere with IgE antibody binding to Der p1 by steric hindrance, but can inhibit the cysteine protease activity of Der p 1. Intra-peritoneal injection of Wan 108, can also prevent and treat mite allergen-induced airway inflammation in sensitized and allergen-challenged mice.44 Hence, anti-protease inhibitor antibodies or other targeted therapies may be used in the future to treat allergic diseases.
THE ROLE OF TLR ACTIVATION AND MOLECULAR MIMICRY IN HDM ALLERGY
The contribution of the TLR4 signaling pathway to HDM allergy was confirmed by the absence of common allergic asthma symptoms in mice deficient in MYD88, TLR4, and IRF3.45,46 Previously, we reported that the differential reactivity of two mouse alveolar macrophage cell lines, MH-S (BALB/c strain, CD14high/TLR4high), and AMJ2-C11 (C57BL/6 strain, CD4low/TLR4low), to Der p could be ascribed to their relative TLR4 expression levels.47 LPS and Der p induced different levels of NO in each cell line. We hypothesize that these different responses depended on the expression of CD14/TLR4 complexes.
In addition, pretreatment of macrophages with SP-D inhibited NO production from Der p and LPS-stimulated alveolar macrophages.47 Recent data also showed that TLR4 expression in lung epithelium, but not on DCs, is essential for the development of a robust HDM-specific Th2 inflammatory response.48 More importantly, TLR4 activation in the airway epithelium by HDM combined with small amounts of LPS induced pro-Th2 cytokines such as TSLP, GM-CSF, IL-25, and IL-33 (Fig. 2). These new findings place epithelial cells in a pivotal position in the development of allergic inflammation though the activation of the TLR4 signaling pathway.
Molecular mimicry is a dominant immunostimulatory effect in the HDM allergen Der p 2. Der p 2 mimics the activity of its fellow ML-domain protein, MD-2, by presenting lipopolysaccharide to TLR4, resulting in the activation of inflammatory genes. In addition, Der p 2 presented with lipopolysaccharide-induced enhanced type 1 allergic sensitization of mice, even when they were deficient in MD-2.49 MD-2 mimicry allows Der p 2 to serve as a self-adjuvant to enhance its allergenicity. Recombinant Der p 2 also stimulates airway smooth muscle cells in a TLR4-independent manner but triggers the MyD88 signal pathway through TLR2.50
We have found that mice repeatedly exposed to mite extracts or the mite component, Der p 2, induced significant NGF production in bronchoalveolar lavage fluid (BALF), activation of AMs and mast cells, and induced eosinophilic infiltration, goblet cell hyperplasia, and hyperplasia of peri-bronchial smooth muscles.51
THE ROLE OF CLR ACTIVATION IN HDM ALLERGY
DCs are the first line of defense in many mucosal membranes. DCs sample their environment using a plethora of receptors, such as C-type lectin receptors (CLRs), scavenger receptors, and TLRs, which increase their internalization efficiency and deliver information regarding the presence of danger signals. DCs also play a role in the induction and re-elicitation of Th2-mediated inflammation in allergic diseases.52,53 However, the molecular processes underpinning these events have remained elusive. In particular, little information is available regarding the mechanism used by DCs to recognize and internalize allergens and how these processes lead to Th2 cell polarization.
The mannose receptor (MR), a C-type lectin, is a multifunctional endocytic receptor on DCs with two distinct lectin activities mediated by its extracellular region.54 The cysteine-rich (CR) domain recognizes sulfated sugars, whereas mannose (together with fucose and N-acetylglucosamine) recognition is mediated by the multiple C-type lectin-like carbohydrate recognition domains (CTLDs). MR mediates internalization of diverse allergens from mite (Der p 1 and Der p 2), dog (Can f 1), cock-roach (Bla g 2), and peanut (Ara h 1) through their carbohydrate moieties.55 Silencing MR expression on monocyte-derived DCs reverses the Th2 cell polarization bias driven by Der p 1 allergen exposure through upregulation of IDO activity.55
Another DC C-type LTR is dendritic cell-specific intercellular adhesion molecule 3-grabbing non-integrin (DC-SIGN). DC-SIGN recognizes carbohydrate structures on pathogens and self-glycoproteins.56 We found that HDM (Der p)-sensitive subjects exhibited decreased expression of DC-SIGN, increased endocytosis, and impaired differentiation of monocyte-derived DCs (MDDCs). Der p allergen may modulate the differentiation and function of MDDCs via interaction with DC-SIGN, a C-type lectin receptor in the mature DCs. Our data indicate that immature MDDCs internalized Der p allergen through DC-SIGN and, after maturation, promoted Th2 polarization of naïve CD4+ T cells (Fig. 3).57
Although Der p 1 can specifically cleave CD40 and DC-SIGN, resulting in a downregulation of Th1 polarization or tolerance through the reduction of IL-12p70 and extracellular thiol production (CD40 cleavage),58,59 our results show that Der p directly binds to DC-SIGN and is pinocytosed into an intracellular compartment in DCs. Downregulation of DC-SIGN on the surface of DCs, which was not due to the protease activity of HDM, will bias DCs towards a Th2 response. DC-SIGN may be important in mediating the innate immune response in DCs when in contact with Der p allergen.
CONCLUSIONS
Type I IgE-mediated hypersensitivity reactions initiated by HDM allergens occur after their recognition by APCs such as dendritic cells (DCs). Recognition leads to Th2 cell differentiation, IgE Ab production, and mast cell sensitization and triggering. Initiation of the allergen response may involve multiple innate immunity receptors on mucosal membranes. Previously, we have demonstrated that extracts from dust mites (Der f) can directly activate innate immune cells, such as alveolar macrophages and mast cells, and can induce Th2 cytokine response without previous in vitro or in vivo sensitization. Recent studies have suggested that allergen-induced DC activation and inflammation may involve TLRs and/or CLRs on antigen-presenting cells.
Using alveolar macrophage cell lines derived from different genetic backgrounds, we found that lipopolysaccharides (LPS) and Der p elicited different responses, and that this was dependent upon the level of expression of the CD14/TLR4 complex. Protease activity in HDM allergens may cause direct damage to the epithelium that leads to production of pro-Th2 cytokines; for example, TSLP, CCL20, IL-25, and IL-33.
The administration of mAb W108, which inhibited IgE binding to the mite allergen epitope, to Der p-sensitize asthma mice alleviated allergen-induced airway inflammation and the reduced the Th2 cytokine immune response. These results suggest a target for the treatment of mite allergen-induced diseases and asthma. Therefore, understanding the role of mite allergen-induced innate immunity may facilitate the development of therapeutic strategies for the treatment of allergic asthma.

 XML Download
XML Download