

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Atopic dermatitis (AD) is a chronic relapsing skin disease that is frequently associated with other atopic conditions such as asthma and allergic rhinitis.1 The skin of AD patients is characterized by immune dysregulation and epidermal barrier defects such as abnormal terminal differentiation of keratinocytes and decreased cornification.2-4 Recent studies showed that these characteristics of AD associate closely with loss-of-function mutations (null) in the filaggrin (FLG) gene. FLG deficiency has been shown experimentally to lead to a failure in the barrier function of the skin in humans.5,6
In mice, loss-of-function FLG mutations result in the absence or reduction of the FLG protein and lead to a compromised skin barrier that allows the entry of allergens, which then trigger an immunological response.7 Several studies also show that FLG null mutations associate significantly with AD. In 2006, Palmer et al.8 reported that the R501X and 2282del4 nonsense mutations were associated with a highly significant dominant risk of AD in many European populations. A study by Ching et al.10 also found that R501X was present in 2.3% of Chinese patients with AD but absent in the controls. Hamada et al.11 found R501X in and two Japanese individuals with severe ichthyosis vulgaris (IV). The E2422X mutation, which was previously found in a single Dutch patient,9 was also detected in one of 100 Singaporean patients with IV12 and two Chinese families with IV, but not in controls.13
The 3321delA mutation appears frequently in Asian populations. It was first reported by a study on 143 Japanese patients with AD: 1.4% of the AD cohort had this mutation while it was absent in Japanese nonatopic and nonichthyotic controls.14 Also, this mutation was reported in Asian including Korean patients with IV.13,15,16 In 2011, Zhang et al.17 showed that the 3321delA was associated with AD and asthma compared to control. However, this mutation was not found in a European population,8 which suggests that this mutation may be more common in the Asian populations.
On the basis of these observations, the present study examined the association between the three FLG mutations 3321delA, R501X, and E2422X and susceptibility to and severity of AD in Korean children.
MATERIALS AND METHODS
Study population
In total, 2,292 children (0-18 years) were enrolled, namely 1,430 patients with AD and 862 normal control subjects who lacked a history of AD or asthma or any atopic manifestations. The subjects with AD were examined at a childhood Asthma Atopy Center, Asan Medical Center in Seoul and diagnosed according to the criteria of Hanifin and Rajka.18 The subjects with AD were divided into two groups: those with positive skin prick test results and specific IgE levels (the allergic AD group) and those with negative results (the non-allergic AD group). AD disease severity was assessed by using the SCORing Atopic Dermatitis index (SCORAD) and the patients with AD were grouped into mild (<15 points), moderate (15-40 points) or severe (>40 points) disease groups. The human ethics committee of the Asan Medical Center Institutional Review Board approved this study. Written informed consent was obtained from the parents of all subjects.
Skin prick test and serum IgE levels
The patients were tested for the 27 common aeroallergens by using the skin prick test. These included the house dust mites Dermatophagoides pteronyssinus, D. farinae, cat, dog, cockroach, the molds Alternaria alternata and Aspergillus fumigatus, grass, tree mix (I), tree mix (II), poak, mugwort, ragweed, egg, and milk etc.19 The allergen-induced wheal that developed after pricking was considered to be positive if it was bigger than the histamine-induced wheal, which was bigger than 3 mm in diameter. Peripheral blood samples were collected from the patients with AD and controls and analyzed for total serum IgE levels and specific IgE levels, including IgE specific for inhalant mix (Dermatophagoides pteronyssinus, Dermatophagoides farinae, cat, dog and cockroach) and food mix (egg, milk, soybean, peanut, fish and wheat). For this, a fluorescent enzyme immunoassay was used (AutoCAP system, Pharmacia Diagnostics AB, Uppsala, Sweden). Antigen-specific IgE concentrations ≥0.35 KIU/L were considered to be positive.
FLG genotyping
Genomic DNA samples were extracted from peripheral whole blood by using the Gentra Puregene Blood kit (QIAGEN Biotech, Seoul, Korea). Genotyping for the FLG mutations 3321delA, E2422X and R501X was conducted by using the TaqMan assay (ABI, Foster City, CA, USA) with a 384-well plate. The final volume of each polymerase chain reaction (PCR) mixture was 5 µL and it contained 10 ng of genomic DNA, 2.5 µL TaqMan Universal PCR Master Mix, and 0.26 µL of 40X Assay Mix.
Statistical analysis
Statistic analyses were performed by using SPSS version 19. The continuous clinical characteristic variables were expressed as mean±standard deviations (SDs) and differences between groups in terms of these variables were tested by using independent t-test. Chi-square test was used to examine differences between groups in terms of categorical variables and to compare the allelic and genotype frequencies. For all analyses, a P value <0.05 was regarded as being statistically significant.
RESULTS
Clinical features of the patient and control groups
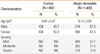
The clinical characteristics of the patients with AD and the controls are presented in Table 1. The patients with AD were 5.17 years old on average and 57.3% were male. The controls were 9.47 years old on average and 49.7% were male. When AD severity was determined by generating objective SCORAD scores, 809, 383, and 210 of the patients with AD were found to have mild, moderate and severe disease, respectively.
Associations between the FLG mutations and AD susceptibility
The patients and controls were genotyped for the three previously reported null mutations 3321delA, R501X, and E2422X, and the association between these mutations and susceptibility to AD was analyzed. The 3321delA deletion was detected in 28 (2.4%) patients with AD but none of the control subjects (P<0.001). Neither E2422X nor R501X were associated with susceptibility to AD: none of the patients with AD and the control subjects had E2422X while only one subject with AD (0.1%) had null mutation of R501X (P=0.446; Table 2). Analysis of the combination of the three null mutations revealed that it associated highly significantly with susceptibility to AD (P<0.001; Table 3)
Associations between 3321delA and AD severity and allergic and non-allergic AD
No significant associations were detected between AD severity and the 3321delA mutation (P=0.842; Table 4). While both the non-allergic AD and allergic AD groups had the 3321delA mutation significantly more frequently than the control group (P<0.001 for both; Table 5), the non-allergic AD and allergic AD groups did not differ in terms of the frequency of this mutation.
DISCUSSION
In the present study, the 3321delA associated significantly with the development of AD. This confirms previous studies that FLG null mutations are associated with AD.10,14,17,20-22 The primary purpose of these study was to identify associations between the FLG mutations R501X, E2422X and 3321delA and susceptibility to and severity of AD in Korean children. In result, we found a significantly higher frequency of the 3321delA and combination of three mutations in AD patients compared to controls. However, we did not find significant association between severity of AD and occurrence of FLG mutation. This finding suggests that FLG mutations may contribute to the genetic susceptibility to AD in Korea as well as in other countries. However, the frequencies of the R501X and E2422X in Koreans were lower than that in Europeans.
Impairment of epidermal barrier function is a clinical hallmark of AD.2-4 FLG is thought to be one of the most important factors in skin barrier function. Mildner et al.5 demonstrated that knockdown of FLG expression in human skin model reproduced the epidermal alterations caused by FLG mutations. Moreover, lack of FLG led to a reduction in the concentration of urocanic acid, which is released by proteolysis of FLG.5 In experimental, immunohistochemical staining for filaggrin revealed that mutation leads to remarkable reduction of filaggrin protein expression in the AD patients' epidermis.23 And the clinical severity significantly correlated with transepidermal water loss, stratum corneum hydration and thickness in filaggrin related AD.24 These findings suggest that FLG dysfunction is an important in the development of AD.
In the last few years, FLG mutations have been identified widely in European, American, and Asian. The previously reported FLG mutations seem to be population-specific: in particular, R501X is common in Europeans9 but is rarely found in Chinese10,17 and Japanese people.11,20 With regard to E2422X and 3321delA were mainly founded in Asian patients with AD.21,22 Thus, three null mutations R501X, E2422X and 3321delA were selected for analysis in the present study. The 3321delA deletion was detected in 28 AD patients (2.4%), but not in controls; and, E2422X and R501X did not associate significantly with susceptibility to AD. The combination of the three mutations was present in 29 (2.6%) patients with AD and none of the healthy controls, indicating a strong association between these FLG mutations and AD (P<0.001; Table 3). These observations indicate that FLG mutations are significant predisposing factors for AD in Koreans.
Overall, the participants in the present study had a low frequency of FLG mutations (ranging from 0% to 2.6%). Similarly, in Chinese patients with AD, only 2.3% and 0.3% had the R501X10 and E2422X22 mutations, respectively. Indeed, the most common mutation in Asian populations is 3321delA, yet it is present in only 9.7% of Chinese patients with AD22 and 1.4% of Japanese patients with AD.14 These results indicate that the frequency of FLG mutations is lower in Asians, including in Korean children, than in Europeans.
The present study also investigated the relationship between FLG mutations and atopy by measuring the frequency of the 3321delA mutation in patients with allergic AD and non-allergic AD. However, the two groups did not differ significantly. This suggests that FLG mutations may have two pathogenic effects. First, FLG mutations may directly affect the structure of the skin itself, thereby reducing its ability to act as a barrier. Supporting this are the associations between FLG mutations and IV in Chinese12,13 and Japanese populations.11 Second, FLG mutation is functionally related to the skin barrier and therefore is most likely to be associated with the cause of skin disease including AD. However, FLG may also affect other allergic disease such as asthma. Supporting this is that FLG variants increase the risk of atopic asthma in the absence of AD or a history thereof,25 and asthma without eczema.26 Also, Chinese patients with AD were interested significant association between FLG mutation and AD with asthma.17 These associations between FLG mutations and allergic disease have been explained by the outside-inside hypothesis.27 It proposes that a barrier abnormality is the driving force behind the development of an inflammatory response in patients with AD. They subsequently hypothesized that reduced levels of FLG, in particular its acidic derivative urocanic acid, increase the pH of the stratum corneum, thereby altering the activity of the multiple serine proteases and two ceramide-generating enzymes that regulate the homeostasis of the stratum corneum.3 Another important downstream consequence of the increased pH and serine protease activity is the generation of active primary cytokines such as IL-1α, IL-1β, IL-25, and IL-33 from their inactive proproteins.28,29 Moreover, defective barrier that leads to a Th2-dominant infiltrate is proposed to be a secondary cause of the inflammation by antigens seen in patients with AD.29 In the present study, FLG mutations were not associated with the severity of AD. Two studies were reported that there was no association between FLG mutation and the severity of AD in Chinese17 and Japanese24 like out results, on the other hand there was a significant association in Singaporean21 and Chinese.10 Also there were several studies that FLG mutations affect to the severity of AD in European.30,31 From these results, the severity of AD may be influenced by FLG mutation in many populations but it seems not to be specific to ethnic. Taken together, FLG is not only a major gene for the development of AD, but that it may also participate in susceptibility to allergic disease. The occurrence of AD in lacking FLG null mutations may be caused by variants in genes pathway that modify filaggrin protein processing or unknown mutation of other independent genes in the skin barrier including stratum corneum and tight junction through environment variations. It is expected that follow-up genetic studies will uncover further genetic loci important in AD.
The present study had several limitations. First, rather than fully sequencing FLG to identify target mutations, three known mutations were examined. Therefore, the effect of other mutations on the development of or severity of AD could not be determined. Second, the present study could not show whether FLG mutation actually causes skin barrier dysfunction. Therefore, additional experiments should be performed to elucidate how FLG mutations promote the development of AD.
In conclusion, FLG null mutations significantly associated with the development of AD in Korean children, although the frequencies of the FLG mutations that were examined were low overall. Mechanism between FLG mutations and Th2 response through defects of skin barrier should be examined for new therapeutic target of AD.




 XML Download
XML Download