

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a chronic inflammatory disease of the airways characterized by airway hyperresponsiveness (AHR) and varying degrees of airflow limitation accompanied by cough, dyspnea, and wheezing. These symptoms are often reversible either spontaneously or with treatment.1 Although most patients' asthma can be controlled with inhaled corticosteroids, some patients are refractory to treatments due to airway remodeling. The histological characteristics of airway remodeling include epithelial denudation, hyperplasia of goblet cells and mucus glands, subepithelial fibrosis, and hypertrophy and hyperplasia of airway smooth muscle. Airway remodeling is thought to be a result of chronic inflammation and is induced by various inflammatory and airway structural cells, in addition to chemical mediators.1-4
Although many studies have suggested that inhaled corticosteroids suppress chronic airway inflammation and airway remodeling, this remains unclear.4 Previous studies have shown that airway remodeling occurs in parallel with inflammation in pediatric patients with asthma and suggests limitations of inhaled corticosteroids.5 For these reasons, therapeutic agents other than corticosteroids, such as cytokine inhibitors, have been developed to suppress airway remodeling.
Asthma pathogenesis is primarily related to T helper type 2 (Th2) cells and their associated cytokines, namely, interleukin (IL)-4, IL-5, and IL-13. Th17 cells and regulatory T (Treg) cells, as well as their cytokines, are correlated with the development of asthma. We previously reported that IL-33 induces Th17-mediated airway inflammation via mast cells in ovalbumin (OVA)-challenged mice.6 Asthma is thought to develop due to an imbalance between Th17 and Treg cells.7,8 Many studies have explored therapeutic agents for asthma such as antibodies (Ab) to IL-5 and IL-4/IL-13 and tumor necrosis factor (TNF)-α inhibitors.8-12 However, anti-IL-5 Abs are only effective in patients with steroid-dependent or steroid-resistant eosinophilic asthma, and TNF-α inhibitors sometimes produce adverse drug effects. Because of this variation in efficacy and the potential for deleterious effects, further studies are required to develop novel therapeutic agents.
IL-9, which was first described in 1980, triggers mast cells and lymphocytes to release chemical mediators and accelerate cell growth and survival. IL-9 is thought to be a Th2 cytokine because of its ability to induce the Th2-biased immune response.13 When Th2 cells or naïve T cells are simultaneously exposed to IL-4 and transforming growth factor (TGF)-β, they undergo phenotype switching driven by transcription factor PU.1 and interferon regulatory factor 4 (IRF4) into IL-9-producing cells (Th9 cells).14 Populations of Th cells other than Th9 cells are also known to produce IL-9 in addition to their own cytokines. Th2 cells produce IL-4, IL-5, and IL-13 in addition to IL-9, and Th17 cells produce interferon (IFN)-γ, IL-17, and IL-9. Although mouse Th9 cells produce both IL-9 and IL-10, human Th9 cells only produce IL-9.15,16 Th9 cells also accelerate chemokine production in airway smooth muscles and epithelial cells, both of which are closely associated with airway remodeling. Previous studies have revealed a close correlation between IL-9 and AHR,17-19 but the relationship between IL-9 and the induction of Th1 or Th17 subsets in allergic asthma remains unclear. In the present study, we examined the relationships between IL-9 and chronic asthma inflammation and investigated whether IL-9 blockade has significant effects on the induction of IL-17-producing cells and airway remodeling in a mouse model of asthma.
MATERIALS AND METHODS
Mice
Eight-week-old female Balb/c mice and C57BL/6 mice were purchased from Daehan Biolink (Eumseong, Korea). Caspase-1 knockout (KO) mice were generously provided by Dr. Sutterwala from the University of Iowa (Iowa City, IA, USA). Mice were housed in the animal care facility with food and water available ad libitum. Mice were exposed to a 12:12 hours light:dark cycle in room air at room temperature (RT). All procedures were approved by the Animal Care and Use Committee of the Ewha Womans University School of Medicine (ESM 11-0164).
Acute allergic asthma model
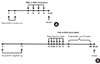
Mice (n=15) were immunized via intraperitoneal injection on days 0 and 5 with 10 µg OVA Grade II (Sigma, St. Louis, MO, USA) complexes with 1 mg aluminum hydroxide (Sigma) at a volume of 0.2 mL per mouse. Using a nebulizer (Medel, Vlaeberg, South Africa), mice were exposed for 30 minutes per day to an aerosol of either 1% OVA in phosphate-buffered saline (PBS) or PBS only on days 12-15 post-immunization. Mice were sacrificed 24 hours after the final aerosol challenge to collect bronchoalveolar lavage fluid (BALF) and lung tissues.
Chronic allergic asthma model
Mice (n=10) were immunized via intraperitoneal injection on days 0 and 12 with 10 µg OVA complex with 1 mg aluminum hydroxide at a volume of 0.2 mL per mouse. Using a nebulizer, mice were exposed to 1% OVA in PBS on days 18-23 for 30 minutes per day. After day 26, mice were exposed to aerosolized 1% OVA three times per week for 5 weeks. Anti-IL-9 Ab (n=5) or control isotype immunoglobulin G (Iso IgG) (n=5) were administered intraperitoneally (100 µg/mouse) once a week during the OVA challenge. A schematic of this protocol is shown in Fig. 1.
BAL Procedure
After the mice were anesthetized with a mixture of Zoletil® (30 mg/kg, Virbac Laboratories, France) and Rompun® (10 mg/kg, Bayer AG, Germany), the tracheas were cannulated. BAL was subsequently conducted with the instillation of 4×0.5 mL PBS into the lungs, along with gentle massaging to maximize cell recovery. BAL cells were pelleted via centrifugation and counted with a hemocytometer. Differential leukocyte counts were obtained on cytospin slide preparations after Giemsa staining.
Histopathology
Lung tissues were fixed in 4% paraformaldehyde and embedded in paraffin. Fixed tissues were cut into 5 µm sections, placed on glass slides, and de-paraffinized. Sections were stained with hematoxylin and eosin (H&E) to detect cellular infiltration, with periodic acid-Schiff (PAS) to detect mucus deposition, and with Sirius Red stain to observe collagen accumulation. The degree of peripheral inflammatory cell infiltration was rated on a 4-point scale: 0, normal; 1, fewer cells, 2, a ring of inflammatory cells up to four cells deep; and 3, a ring of inflammatory cells more than four cells deep.20 Tissue sections from each mouse were measured using image analysis software (analySIS FIVE, Olympus Corporation, Japan) to analyze the deposition of mucus and collagen.
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assays (ELISAs) were performed to measure IFN-γ, IL-4, and IL-5 with BD OptEIA™ (BD Biosciences, San Diego, CA, USA). IL-9, IL-17, and IgE concentrations in BALF were measured using a commercially available kit from BioLegend (San Diego, CA, USA). Wells were coated with 100 µL capture Ab in a coating buffer (0.1 M sodium carbonate, pH 9.5) and plates were incubated overnight at 4℃. Wells were rinsed with a washing buffer (PBS with 0.05% Tween-20) and reactions were stopped with assay diluent (10% fetal bovine serum in PBS) for 1 hour at RT. After the supernatant was obtained by BALF centrifugation, 100 µL supernatant was put in each well and incubated for 2 hours at RT. The supernatant was removed, the wells were washed, and 100 µL detection Ab and streptavidin-conjugated horse radish peroxidase (HRP) reagent was added and reacted for 1 hour at RT. After the reaction, the wells were washed, 100 µL substance solution (tetramethylbenzidine and hydrogen peroxide, BD Biosciences) was added to each well, and the plates were incubated in the dark for 30 minutes at RT. The absorbance was measured at 450 nm within 30 minutes of adding the stop solution (2N H2SO4).
Flow cytometry
To compare cellular profiles after the induction of airway inflammation, BAL cells from mice challenged with OVA with or without anti-IL-9 Ab were stained and analyzed on a FACSCalibur system (Becton Dickinson, Franklin Lakes, NJ, USA). The Abs used for flow cytometry were as follows: FITC-anti-mouse CD49d (9C10, BD Biosciences), FITC-anti-mouse Ly-6G/Ly-6C (Gr-1; RB6-8C5, BioLegend), PE-anti-mouse FcεRIα (MAR-1, BioLegend), APC-anti-mouse CD117 (c-kit; ACK2, BioLegend), FITC-anti-mouse CD49b (HMα2, BioLegend), FITC-anti-mouse CD19 (6D5, BioLegend), anti-mouse CD4 (GK1.5, BioLegend), APC-anti-mouse IFN-γ (XGM1.2, BioLegend), anti-mouse IL-4 (11B11, BD Bioscience), anti-mouse IL-9 (RM9A4, BioLegend), anti-mouse IL-17A (TC11-18H10.1, BioLegend), and Alexa Fluor®488-anti-mouse FoxP3 (206D, BioLegend).
Immunoblot
BAL cells were lysed in a lysis buffer (1% Triton X-100, 150 mM NaCl, 20 mM Tris, pH 7.5) with protease inhibitors, and 10 µg protein was separated using 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The resolved proteins were transferred to polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). Membranes were blocked with 5% skim milk in Tris-buffered saline and incubated with the following primary Abs: anti-IL-1β (#2022, Cell Signaling Technology, Danvers, MA, USA), anti-β-actin (AC-15, mouse monoclonal, Abcam, Cambridge, UK), and anti-PU.1 (#2266, Cell Signaling Technology). After washing, the membranes were incubated with HRP-conjugated anti-mouse or anti-rabbit IgG secondary Ab, and the signals were visualized using an enhanced chemiluminescence kit (Thermo Fisher Scientific Inc., Waltham, MA, USA), in accordance with the manufacturer's instructions.
RESULTS
OVA-induced airway inflammation
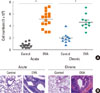
We used OVA in a standard sensitization protocol in Balb/c mice to induce acute (Fig. 1A) or chronic airway inflammation (Fig. 1B). BAL was performed on the mice 24 hours after the last challenge via the inhalation of 1% OVA aerosol solution to determine the airway inflammation state. We compared total BAL cells and pathological changes after OVA inhalation because inflammatory pulmonary disease is characterized by the infiltration of inflammatory cells into the airway and tissue. BALF from OVA-challenged Balb/c mice exhibited higher total cell numbers, regardless of the duration of OVA challenge (Fig. 2A). In acute asthma inflammation, the total numbers of inflammatory cells were 0.71±0.39×106 cells/mL in the control group and 5.13±1.35×106 cells/mL in the acute asthma group. In chronic asthma inflammation, there were 1.95±0.86×106 cells/mL in the control group and 4.58±1.21×106 cells/mL in the chronic asthma group (Fig. 2A). H&E-stained lung tissue specimens showed more inflammatory cellular infiltrates in both the acute and chronic asthma groups than in the control groups (Fig. 2B).
Cytokine secretion upon airway inflammation
Inflammation associated with asthma is regulated by multiple pro-inflammatory mediators. We measured the concentrations of IFN-γ, IL-4, IL-5, IL-9, IL-17, and IgE to compare the type of immune response elicited by different durations of OVA challenge. IL-4, IL-5, and IgE concentrations were significantly higher in the acute and chronic asthma groups than in either of the control groups. In the acute asthma group, IL-4 increased from 18.17±2.36 pg/mL to 112.67±21.65 pg/mL, IL-5 increased from 9.33±1.89 pg/mL to 143.57±27.67 pg/mL, and IgE increased from 0.04±0.02 ng/mL to 6.69±0.76 ng/mL. In the chronic asthma group, IL-4 increased from 19.00±1.00 pg/mL to 44.33±5.01 pg/mL, IL-5 increased from 10.20±2.80 pg/ml to 86.00±8.72 pg/mL, and IgE increased from 0.04±0.02 ng/mL to 4.17±0.51 ng/mL (Fig. 3).
IFN-γ, IL-9, and IL-17 concentrations only increased in the chronic asthma group. There were some differences in the concentration of IFN-γ (25.33±9.61 pg/mL in acute asthma vs. 41.00±13.08 pg/mL in controls), IL-9 (65.33±5.03 pg/mL in acute asthma vs. 107.33±11.02 pg/mL in controls), and IL-17 (32.43±2.10 pg/mL in acute asthma vs. 36.97±3.94 pg/mL in controls), but the differences were not statistically significant. However, in the chronic asthma group, IFN-γ increased from 21.67±2.89 pg/mL to 95.67±14.64 pg/mL, IL-9 increased from 60.67±9.24 pg/mL to 588.00±73.73 pg/mL, and IL-17 markedly increased from 35.87±4.94 pg/mL to 101.33±17.04 pg/mL (Fig. 3). These results indicate that airway inflammation induced by OVA aerosol challenge results in prominent Th2 responses in acute asthma mice, whereas chronic asthma mice display heterogeneous responses with a significant increase in IL-17 and IL-9 secretion in the lung.
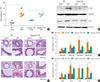
Caspase-1 KO mice exhibit lower numbers of IL-17-producing cells and induced airway inflammation
During the development of allergic airway inflammation, caspase-1 activation via the inflammasome pathway leads to the production of the active form of IL-1β and contributes to the enhancement of Th17 cell differentiation.21 We induced chronic airway inflammation in caspase-1 KO mice to investigate the role of Th17 cells in asthma because caspase-1-mediated production of IL-17 is correlated with disease severity in asthmatic patients.22 We found that total BAL cells counts were increased in OVA-challenged caspase-1 KO mice (Fig. 4A), and IL-1β expression was primarily increased in wild-type OVA-challenged (WT-OVA) mice (Fig. 4B). PU.1 expression was increased in both WT-OVA and caspase-1 KO mice (Fig. 4B). Although WT-OVA mice and caspase-1 KO mice both showed increased recruitment of inflammatory cells and mucus secretion (Fig. 4C), the only difference in cell numbers between the WT and caspase-1 KO mice was observed in IL-17-producing CD4+ cells. We confirmed that a decrease in IL-1β affected Th17 subsets but not IL-9-producing CD4+ cells. Moreover, we observed that an increase in IL-9-producing CD4+ cells did not affect the induction of Th17 cells in OVA-challenged caspase-1 KO mice (Fig. 4D and E).
IL-9 blockade suppresses pulmonary inflammation
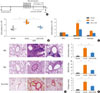
IL-9 expression is elevated in lungs of asthmatic patients, and IL-9 receptor (IL-9R) expression can be observed in the lungs of asthmatic individuals but not healthy controls.23-25 IL-9 has biological effects on a number of distinct cell types, and may have other effects on immune cells and resident tissue cells that contribute to the development of inflammation beyond its first description as a T-cell or mast cell growth factor. Our ELISA (Fig. 3) results suggest that IL-9 may be a crucial inflammatory mediator in chronic airway inflammation, and is not affected by IL-17 in caspase-1 KO mice (Fig. 4E). We utilized a model of chronic asthma inflammation in which OVA-sensitized Balb/c mice were subjected to a 5-week exposure to OVA to investigate the effect of anti-IL-9 Ab treatment on chronic inflammation and airway remodeling (Fig. 5A).
Mice were sacrificed after the last aerosol challenge, BALF was collected, and different cell types were counted to assess the effect of anti-IL-9 Ab on pulmonary inflammation. The total numbers of inflammatory cells in BALF were (2.54±0.73)×106 cells/mL in the untreated (control) group, (4.94±0.48)×106 cells/mL in the Iso Ab group, and (3.20±0.32)×106 cells/mL in the anti-IL-9 Ab group. The difference in cell numbers observed between the Iso Ab group and the anti-IL-9 Ab group was statistically significant (Fig. 5B). Specifically, there were significant changes in the number of eosinophils. The numbers of eosinophils were (0.25±0.07)×106 cells/mL in the control group, (3.75±0.35)×106 cells/mL in the Iso Ab group, and (1.95±0.35)×106 cells/mL in the anti-IL-9 Ab group. Monocytes and basophil cell counts decreased without statistical significance (Fig. 5C).
Histological examination of the lung tissue revealed that chronic OVA challenge induced extensive infiltration of inflammatory cells in peri-vascular and peri-bronchial tissues. This infiltrate was severe in the Iso Ab group, but markedly depressed in the anti-IL-9 Ab group. The inflammation indices, described in the Materials and Methods, were 0.25±0.5 in the control group, 2.75±0.5 in the Iso Ab group, and 1.25±0.5 in the anti-IL-9 Ab group. The mucus indices, measured by PAS staining, were 1.33%±0.15% in the control group, 13.63%±1.37% in the Iso Ab group, and 5.60%±0.82% in the anti-IL-9 Ab group (*P<0.05). The collagen indices, as calculated from the results of Sirius Red staining, were 0.46%±0.12% in the control group, 3.40%±0.82% in the Iso Ab group, and 1.77%±0.35% in the anti-IL-9 Ab group (*P<0.05; Fig. 5D and E). Together, these results suggest that neutralizing IL-9 is effective for preventing or halting tissue fibrosis during chronic asthmatic responses.
IL-9 blockade affects cellular profiles in chronic airway inflammation
To determine the profile of airway-infiltrating cells in OVA-challenged mice with or without anti-IL-9 Ab, we analyzed changes in inflammatory cells such as eosinophils, neutrophils, mast cells, basophils, and B cells in BALF by flow cytometry (Fig. 6A). The proportion of eosinophils decreased from 34.33%±3.51% to 21.67%±3.51% in the anti-IL-9 Ab group and correlated well with the results shown in Fig. 5C. Neutrophils decreased from 37.33%±2.51% to 16.33%±1.53%, mast cells decreased from 22.67%±2.52% to 10.00%±2.00%, basophils decreased from 13.33%±3.06% to 8.33%±1.53%, and B cells decreased from 32.33%±3.21% to 22.33%±2.52%. These differences were statistically significant (Fig. 6B). Next, we examined the proportion of subsets of CD4+ helper T cells to compare the lymphocytic immune response after administration of anti-IL-9 Ab (Fig. 6C). In the anti-IL-9 Ab group, the proportion of IL-9-producing T cells significantly decreased from 23.00%±3.00% to 9.00%±2.00%, and the proportion of IL-17-producing cells significantly decreased from 12.67%±2.52% to 6.00%±1.00%. The proportions of IL-4-producing T cells and Foxp3-positive T cells did not change significantly. The proportions of IFN-γ-producing CD4+ T cells were 0.72%±0.37% in the control group, 3.33%±1.53% in the Iso Ab group, and 0.36%±0.21% in the anti-IL-9 Ab group (Fig. 6D).
IL-9 blockade alters the production of cytokines and IgE during chronic OVA challenge
Next, we evaluated the effect of anti-IL-9 Ab treatment on IgE and cytokines in the lung (Fig. 7). IL-4 and IFN-γ were markedly increased in the Iso Ab group compared to the control group, but the decrease after IL-9-neutralizing Ab injection was not statistically significant. IFN-γ levels were 29.69±7.37 pg/mL in the control, 62.5±12.02 pg/mL in the Iso Ab group, and 49.67±13.65 pg/mL in the anti-IL-4 Ab group. IL-4 levels were 20.67±2.89 pg/mL in the control group, 37.67±3.79 pg/mL in the Iso Ab group, and 31.33±3.21 pg/mL in the anti-IL-9 Ab group. In contrast, IL-5, IL-9, IL-17, and IgE levels were significantly lower in the anti-IL-9 Ab group than in the Iso Ab group. IL-5 levels were 31.00±3.61 pg/mL in the control group, 75.00±6.24 pg/mL in the Iso Ab group, and 47.67±1.53 pg/mL in the anti-IL-9 Ab group. After IL-9 Ab administration, IL-9 levels were 67.67±34.27 pg/mL in the control group, 759.33±179.05 pg/mL in the Iso Ab group, and 347.33±24.79 pg/mL in the anti-IL-9 Ab group. IL-17 levels were 13.33±1.15 pg/mL in the control group, 58.33±16.17 pg/mL in the Iso Ab group, and 28.33±9.29 pg/mL in the anti-IL-9 Ab group. IgE levels were 0.02±0.00 ng/mL in the control group, 4.20±0.28 ng/mL in the Iso Ab group, and 2.20±0.57 ng/mL in the anti-IL-9 Ab group (Fig. 7). Based on these results, IL-9 blockade significantly attenuated the levels of IL-5, IL-9, IL-17, and IgE. These data indicate that the suppression of pulmonary inflammation by anti-IL-9 Ab is mediated through a decrease in multiple inflammatory mediators, including IL-17.
DISCUSSION
We examined the relationship between IL-9 and chronic airway inflammation to determine if IL-9 blockade would affect IL-17-driven airway inflammation. After anti-IL-9 Ab administration, the number of inflammatory cells and Th17 cells in BALF, as well as the inflammation index, mucus index, and the collagen index, were significantly lower in the chronic asthma group.
IL-9 has direct and indirect effects on lymphocytes, mast cells, epithelial cells, smooth muscle cells, and hematopoietic progenitors that affect the development of immunity and inflammation. In asthma patients, IL-9R is found on mast cells and polymorphonuclear cells in the lung.26 Lung-selective expression of IL-9 is known to cause airway inflammation characterized by infiltrating eosinophils and lymphocytes and pathologic changes including epithelial cell hypertrophy, mucus production, and increased subepithelial deposition of collagen.27 Transgenic mice systemically over-expressing IL-9 display an elevated AHR to bronchoconstrictors, enhanced eosinophilic inflammation of the airways, and elevated serum IgE following sensitization and challenge with the mold Aspergillus fumigates.17 However, goblet cell hyperplasia and mastocytosis are severely impaired in IL-9-deficient mice in response to pulmonary challenge.28
IL-9 induces differentiation and proliferation of Th17 cells, enhances immunologic tolerance of Treg cells without proliferation, and increases IgE production in B cells.13 However, our results showed that even though IL-9+ CD4+ T cells were increased in OVA-challenged caspase-1 KO mice, they did not affect the number of IL-17-producing CD4+ T cells (Fig. 4E). However, after treatment with an IL-9 neutralizing Ab, the proportion of Th17 cells and secretion of IL-17 was decreased (Figs. 6D and 7). Our data suggest that IL-9 can promote the development of Th17 cells at sufficient IL-1β concentrations.
There have been several studies on changes in inflammatory reactions induced by anti-IL-9 Ab administration. Cheng et al.29 demonstrated that anti-IL-9 Ab treatment suppresses airway inflammation and hyperreactivity to methacholine challenge. They found that the Th2 cytokines IL-4, IL-5, and IL-13 were decreased in BALF after exposure to anti-IL-9 Ab, which does not agree with our results. This discrepancy may due to the fact that anti-IL-9 Ab was administered during the acute stage of inflammation in the previous study, whereas we administered the Ab during the chronic stage of inflammation. Kearley et al.30 indicated that, in a chronic asthma model, IL-9 blockade reduces the number of mast cells and production of vascular endothelial growth factor, fibroblast growth factor 2, and TGF-β, and thus suppresses AHR and remodeling. Th2 cells play a crucial role in acute allergic reactions, but not during late-phase allergic reactions and airway remodeling.31 It is interesting to note that IL-9 expression increases when Th2 cells are exposed to IL-4 and TGF-β (a representative cytokine for airway remodeling). Soroosh and Doherty32 suggested that some effector Th2 cells may switch to Th9 cells and play a role in distinct aspects of chronic lung inflammation mediated by IL-9 in the presence of IL-4 and TGF-β.
Two randomized phase-IIa clinical trials of a humanized anti-IL-9 monoclonal Ab have been performed. These studies showed that anti-IL-9 Ab has linear pharmacokinetics, induces no Ab to anti-IL-9 Ab, and has a similar frequency of adverse drug effects as the placebo.33 They also described that the maximal therapeutic effect was achieved 56 days after administration of the monoclonal Ab, which suggests that IL-9 blockade may be more effective for treating chronic allergic inflammation rather than acute allergic inflammation. Further studies with a larger sample size and a longer duration of drug administration are required before applying these results to clinical practice. In the same clinical trials, they administered the Ab twice weekly for 8 weeks, which is not feasible in a clinical setting. Safe application of anti-IL-9 Ab in patients with asthma requires longer intervals between individual drug administrations using an effective drug-delivery system.
In conclusion, anti-IL-9 Ab can reduce cellular infiltration related to chronic airway inflammation and remodeling. Treatment with an anti-IL-9 Ab can reduce the number of Th9 cells, Th17 cells, mast cells, eosinophils, and neutrophils in the airway, inhibit the synthesis and secretion of cytokines, and reduce IgE synthesis in B cells. Further studies should be performed to utilize an anti-IL-9 Ab to treat asthma patients because this Ab can improve various pathophysiological changes.



 XML Download
XML Download