

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Nasal polyposis (NP), which is a heterogeneous, chronic, inflammatory disease of the sinuses, is often associated with asthma1,2 and is characterized by a mixed Th1 and Th2 immune response.2,3 Although the precise mechanisms underlying the pathologic mechanisms of NP are not well-defined, nasal polyps have recently been reported to be characterized by the presence of numerous eosinophils, activated mast cells, and Th2-type cells.2-4 In addition, the numbers of Th2-type cells are increased, irrespective of the patient's atopic status. Studies have also demonstrated strong local upregulation of IL-13, thymus and activation-related chemokine (TARC), and immunoglobulin E (IgE) synthesis with the formation of specific IgE to Staphylococcus aureus enterotoxins, suggesting a possible role for superantigens in these pathologic processes.5,6 Recent studies have also suggested that S. aureus secretes exotoxins that act as superantigens and upregulate the variable beta region of lymphocytes in chronic hyperplastic sinusitis with nasal polyposis.7
More recently, we have demonstrated increased expression of thymic stromal lymphopoietin (TSLP) in nasal polyps, irrespective of the atopic status of the patient, as compared to the level of expression in allergic nasal mucosa.8 The level of TSLP expression was in good correlation with the levels of eosinophils and IgE in the nasal polyps, suggesting a crucial role for TSLP in driving the severity of Th2-type inflammation in the nasal polyps and increased eosinophilia.
Forkhead box P3 (FOXP3) plays important roles in the development and function of CD4(+) regulatory T cells (Tregs) and represents a specific marker for these cells. Tregs are important in balancing immune responses and maintaining peripheral tolerance. Defects in Tregs have been implicated in the pathogenesis of chronic inflammatory and autoimmune diseases, such as idiopathic thrombocytopenic purpura, burning mouth syndrome, and allograft rejection, which have decreased expression of FOXP3.9-11 This may reflect the importance of FOXP3 for the maintenance of normal tissues. Current evidence suggests that allergic disease and asthma are also characterized by deficiencies in Tregs, which allow Th2 cells to expand.12-14 Despite the observed increases inTh2-type cells and IgE levels in the nasal polyps, irrespective of the patient's atopic status, there are no studies comparing the numbers of Tregs in nasal polyps and allergic nasal mucosa.
To study the role of FOXP3 in the pathogenesis of nasal polyps, we compared the levels of FOXP3 expression in the nasal polyps and nasal mucosa of patients with allergic rhinitis.
MATERILAS AND METHODS
Seventeen patients with NP (12 males and 5 females; mean age, 30.6 years) were included in this study. Two of the enrolled patients had asthma. Seven of the patients with NP were atopic with perennial allergic rhinitis (PAR) and ten of them were non-atopic. Six of the PAR patients with NP had typical nasal allergy to house dust mite (HDM) and one had nasal allergy to cat and dog dander, as diagnosed using history and the radioallergosorbent test (RAST). Of the six patients with HDM allergy, four also had seasonal allergic rhinitis to Japanese cedar pollen. In the present study, we also included fifteen patients with PAR (13 males and 2 females; mean age, 28.75 years) who had typical clinical symptoms of nasal allergy, comprising sneezing, rhinorrhea and nasal congestion, and positive serum-specific IgE, as analyzed by RAST.
The diagnosis of allergic rhinitis (AR) was based on a history of clinical symptoms of sneezing, rhinorrhea, and nasal congestion, clinical examination by anterior rhinoscopy and, when there was a positive history, a positive serum-specific IgE by RAST. All patients were symptomatic at the time of collecting the specimens and none of them had previously received immunotherapy. Nasal polyp specimens and nasal mucosal tissue samples were collected at surgery performed as part of the treatment either for the removal of nasal polyps or for the resection of hypertrophied turbinates. All medications were prohibited for at least one month prior to surgery. The study was approved by the Nippon Medical School Medical Ethics Committee and informed consent was obtained from all patients.
Collection and processing of specimens
Nasal polyp tissues were obtained at surgery done as a part of the treatment for the removal of nasal polyps (polypectomy/functional endoscopic sinus surgery). Nasal mucosal specimens were obtained at surgery (conchotomy) performed for the treatment of hypertrophied turbinates. Both nasal polyp specimens and nasal mucosal tissue specimens (3×4 mm) were rinsed in phosphate-buffered saline (PBS; pH 7.6), and then processed for immunohistochemistry as described below.
Immunohistochemistry for the detection of FOXP3
The nasal polyp and nasal mucosa tissue specimens were first fixed in periodate lysine paraformaldehyde (PLP), and then incubated for 4 hours each in gradients of sucrose that varied from 10% to 20% in PBS. Subsequently, the specimens were embedded in Tissue Tech OCT compound, frozen in liquid nitrogen, and stored at -80℃ until further use. To investigate the expression of FOXP3 and to localize the protein in the nasal polyps and nasal mucosa, immunohistochemistry was performed.
For immunohistochemistry, frozen sections of PLP-fixed specimens were cut at 5-µm thickness using a cryostat (Sakura Finetek, Tokyo, Japan), air-dried for 30 minutes, and then fixed in acetone for 10 minutes. Immunohistochemistry was performed using the peroxidase-based avidin-biotin complex (ABC) method (Vectastain ABC kit; Vector Laboratories, Burlingame, CA, USA). Briefly, acetone-fixed sections were rehydrated by incubation in Tris-buffered saline (TBS; pH 7.4). Thereafter, the specimens were incubated in 10% H2O2 for 30 minutes to block endogeneous peroxidase. After two rinses in TBS, the specimens were incubated for 45 minutes in Protein Block Serum Free Ready to Use (Dako, Glostrup, Denmark). After two rinses in TBS, the sections were incubated overnight at 4℃ in the optimal concentration of primary mouse monoclonal antibody against FOXP3 (5 µg/mL) (clone 236A/E7, sc-56680, lot # K2707; Santa Cruz Biotechnology, Santa Cruz, CA, USA). IHC staining was then performed using the HRP method according to the manufacturer's instructions. After rinsing in TBS that contained 0.1% Tween-20 (TBS-t; pH 7.6), the sections were incubated in the ABC. The sections were then rinsed in TBS-t. Specific immunoreactivity of the anti-FOXP3 antibodies was visualized by incubating the tissues in multiple-adsorption biotinylated secondary horse anti-mouse IgG antibody (1:200 dilution, Vectastain Elite ABC Reagent; Vector Laboratories) and the 3-amino-9-ethyl carbazole (AEC) substrate chromogen (Ready to Use kit; Dako). All rinses were carried out in TBS-t (pH 7.6) at room temperature. The reaction was stopped by immersing in distilled water. The sections were counterstained with Mayer's hematoxylin and mounted in glycergel mounting medium (C0563; Dako). As a negative control, the primary antibodies were substituted with an irrelevant isotype-matched mouse IgG (IgG1; Dako).
Double-immunohistochemistry for the detection of FOXP3+CD4+ cells
Although FOXP3 is a biomarker of Tregs, it also expressed in some non-CD4+ cells.16 To confirm that the FOXP3+ cells are Tregs, we assessed FOXP3 and CD4 expression by double-immunohistochemistry using the labeled streptavidin biotin (LSAB) and alkaline phosphatase methods, so as to identify FOXP3+ CD4+ cells.15 Processing of the tissue samples for double-immunohistochemistry was performed as previously described for single staining for FOXP3. For double-immunohistochemistry, we performed the first immunostaining with HRP, as in the previous single immunohistochemistry, using HRP label detection with 3,3'-diaminobenzidine (DAB-brown) for the first primary FOXP3 mouse mAb (clone 236/E7; AbD Serotec, Kidlington, UK). Subsequently, the tissue sections were incubated in an elution buffer of 10 mM citrate and 0.05% Tween-20 (pH6.0), which had been heated to 60℃ for 10 minutes before the second immunohistochemistry, which involved the alkaline phosphatase-labeled detection system with Fast Red Substrate Systems (Dako) for the anti-CD4 mouse mAb (clone MT 310, sc-19641; Santa Cruz Biotechnology). For the negative control, the second primary antibodies were substituted with an irrelevant isotype-matched mouse IgG (IgG1) (Dako).
Cell counting
The numbers of positively stained FOXP3+ cells and FoxP3+ CD4+ cells were counted under an Olympus microscope in six randomly selected visual fields using an objective micrometer in an area of 0.202 mm2 and at a magnification of 400 HPF. The average for the six fields was taken as the cell count for that section.
Epithelia and lamina propria were evaluated separately. The epithelium, defined as the area between the cilia and basal membrane, was evaluated by counting positive cells per 0.0625 mm2. The lamina propria, defined as the area immediately underneath the basal membrane, was evaluated by counting positively stained cells in six randomly selected areas per high-power field (area: one high-power field at 400× is equivalent to 0.0625 mm2). The results are expressed as the number of cells per 0.0625 mm2 in the epithelial and subepithelial areas and compared between the nasal mucosa and nasal polyp tissues.
Statistical analysis
The data are presented as mean±SD. Statistical analyses were carried out using the Student's t-test. Differences were considered statistically significant only when the P-values were less than 0.05. All statistical analyses were performed using the SPSS ver. 15 software (SPSS, Chicago, IL, USA).
RESULTS
Immunolocalization of FOXP3+CD4+ cells in nasal polyps and allergic nasal mucosa
A large proportion of the CD4+ cells expressed FOXP3 (Fig. 3A and B). Almost all the FOXP3+ cells were CD4+. The negative control (isotype-matched IgGs) did not show immunoreactivity for either FOXP3 or CD4 (Fig. 3C).
Comparison of the numbers of FOXP3+ cells in the epithelia and lamina propria of nasal polyps
The number of FOXP3+ cells in the epithelia of the nasal polyps group was 2.39±2.33 cells per 0.0625 mm2. In the lamina propria, the number of FOXP3+ cells was 2.79±1.74 per 0.0625 mm2 (Fig. 4A). No statistically significant difference was detected for the number of FOXP3+ cells between the epithelia and lamina propria.
Comparison of the numbers of FOXP3+ cells in the epithelia and lamina propria of the nasal mucosa of patients with allergic rhinitis
The number of FOXP3+ cells in the epithelia of the nasal mucosa of allergic patients was 3.60±2.64 cells per 0.0625 mm2. The number of FOXP3+ cells in the lamina propria of the nasal mucosa of allergic patients (5.99±4.29 cells) was significantly higher than that in the epithelial layer (P=0.009) (Fig. 4B).
Comparison of numbers of FOXP3+ cells in nasal polyps and AR nasal mucosa
The numbers of FOXP3+ cells were significantly lower in the lamina propria of the nasal polyps than in the lamina propria of the nasal mucosa of the AR patients (2.79±1.74 vs. 5.99±4.29, P=0.008) (Fig. 5A). Although the number of FOXP3+ cells was higher in the epithelia of the allergic nasal mucosa, as compared to that in the nasal polyps (3.60±2.64 vs. 2.39±2.33), the difference was not statistically significant (Fig. 5B).
Comparison of the number of FOXP3+CD4+ cells in nasal polyps from atopics and non-atopics
There was no difference in the numbers of FOXP3+ cells in the nasal polyps obtained from atopics and non-atopics, with respect to both the epithelia (2.70±1.90 vs. 2.07±2.71) and lamina propria (3.52±2.04 vs. 2.25±1.41).
Comparison of the numbers of FOXP3+CD4+ cells in nasal polyps and allergic nasal mucosa.
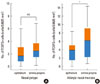
Double immunohistochemistry for FOXP3 and CD4 was performed to detect cells that stained positively for both FOXP3 and CD4. The number of FOXP3+CD4+ cells was significantly higher in the nasal mucosa of the AR patients than in the nasal polyps (84.83%±5.73% vs. 63.18%±15.59%, P=0.002) (Fig. 6).
DISCUSSION
Tregs represent a subset of CD4+ cells that suppresses the functions of other lymphocytes and are characterized by surface expression of CD4 and CD25 and nuclear expression of the transcription factor FOXP3. Recently, many functions of FOXP3 and its influence on the immune system have been elucidated. Tregs influence the development and expression of atopy and the allergic response.9-11 Adaptive FOXP3+ Tregs are essential for establishing mucosal tolerance and for suppressing IL-4 production.17 Decreased expression of FOXP3 has been reported in asthma and allergic rhinitis,12,18 although there have been no studies to compare FOXP3 expression in nasal polyps and allergic nasal mucosa or to define the role of Tregs in NP pathogenesis. In the present study, we compared the levels of FOXP3 expression in allergic nasal mucosa and nasal polyps and demonstrated lower expression of FOXP3 in nasal polyps.
Recently, NPs were characterized by a polarized Th2 reaction and eosinophilic inflammation and high IgE levels, irrespective of atopic status. We have recently reported that high-level expression of TSLP correlates strongly with the numbers of eosinophils in nasal polyps and also IgE, irrespective of atopic status.8 In the present study, we demonstrate decreased numbers of FOXP3+ cells in nasal polyps, as compared to allergic nasal mucosa. These lower numbers of FOXP3+ cells observed in patients with NP may result in unopposed overexpression of Th2 cells and the increased migration and activation of dendritic cells, eosinophils, and mast cells.
High IgE concentrations have been found in nasal polyps, indicating that IgE is produced locally.19-21 Suh et al.20 also reported a polyclonal hyper-immunoglobulinemia E, associated with the presence of IgE specific for S. aureus enterotoxins (SAE), colonization with S. aureus, and increased eosinophilic inflammation in a relevant subgroup (about 50%) of nasal polyp patients. These authors also reported a strong correlation between the levels of IgE in nasal polyps and the presence of nasal eosinophils. Our recent study demonstrates that the high levels of IgE seen in nasal polyps are in good correlation with the number of TSLP+ cells in nasal polyps as well as between TSLP and eosinophils, irrespective of atopic status.8 Moreover, the levels of TARC/CCL17 and macrophage-derived chemokine (MDC) were also high in the nasal polyps.8 In the present study, we also found lower numbers of CD4+FOXP3+ cells in the nasal polyps than in the allergic nasal mucosa, irrespective of atopic status.
It has been suggested that S. aureus secretes exotoxins that may act as superantigens and upregulate the variable beta region of lymphocytes in chronic hyperplastic sinusitis with NP.7 Recently, Van Bruaene et al.22 reported lower expression of FOXP3 mRNA in chronic rhinosinusitis. In the present study, we compared the levels of expression of FOXP3 in the nasal mucosa of patients with AR and NP, as well as in the nasal polyps from atopics and non-atopics. While the numbers of Tregs in the nasal mucosa of patients with AR are reported to be low, we detected a decrease in the number of CD4+FOXP3+ Tregs in both the atopic and non-atopic nasal polyps. In this context, Pérez Novo et al.23 recently assessed the effect of S. aureus enterotoxin B (SEB) on T-cell activation in patients with nasal polyps and asthma, and its possible link to aspirin hypersensitivity. SEB significantly increased the levels of IFN-γ, IL-4, TNF-α, IL-5, and IL-2 in the supernatants of both NP polyp groups with asthma but with and without aspirin sensitivity compared with controls. The baseline level of FOXP3 was significantly decreased in both NP-asthma groups. After incubation with SEB, FOXP3 was significantly up-regulated in the control group, but not in the NP-asthma groups. These researchers concluded that although SEB induces both Th1 and Th2 pro-inflammatory responses in patients with NP and asthma, regardless of the presence of aspirin hypersensitivity, the nature of this response may be linked to the basal deficiency of FOXP3 observed in the NP-asthma patients. Therefore, the deficiency of FOXP3 may lead to the enhanced Th2 inflammation in the NP of atopics and non-atopics and the high level of IgE, irrespective of the atopic status.
These data suggest that the suppressive function of Tregs may be partially inhibited. This might partially explain the more pronounced Th2-type inflammation and eosinophilic inflammation in nasal polyps, as compared to the allergic nasal mucosa. Furthermore, there is no reported statistically significant difference between the lymphocyte subpopulations of atopic and non-atopic patients.24,25
Taken together, the present findings indicate that the deficiency of Tregs in nasal polyps may play an important role in enhancement of the severity of Th2 inflammation in nasal polyps and the persistence and progression of NP. Furthermore, these results point to increasingly important roles for T cells in chronic rhinosinusitis with nasal polyposis.





 XML Download
XML Download