

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Both inflammation and structural changes may occur in allergic rhinitis (AR), and chronic eosinophilic inflammation is related to the development of remodeling.1 Remodeling is characterized by epithelial fragility, reticular basement membrane thickening, and an increase in vascularity, the number of fibroblasts, and interstitial collagen as well as mucous gland hypertrophy.2 Remodeling appears to be less extensive in the nose than in the lower airways because of the differences in the secretory activity of smooth muscle cells that are present in the bronchi but not in the nose. The different embryologic origins of the bronchi and nose may also account for the differences in the extent of remodeling between different airway regions.3 Knowledge of fibrogenic growth factors in the nasal mucosa of AR patients is lacking because of the paucity of studies on AR compared to asthma.4 Periostin (formerly called osteoblast-specific factor 2) was originally identified as a secreted factor in a screen of a mouse osteoblastic library. It is thought to function in bone adhesion, because its sequence is similar to that of an insect adhesion protein, fasciclin-1, and because it is expressed in specialized connective tissues that form and support mineralized tissue.5 This protein is induced by interleukin (IL)-4 and IL-13 in airway epithelial cells.6,7 Periostin also shares structural homology to other mammalian proteins, including big-h3, stabilin I and II, MBP-70, algal-cell adhesion molecule, and periostin-like factor.8-12 To date, many studies on periostin have focused on its role in extracellular matrix deposition, fibrosis, and tissue remodeling following myocardial infarction, as well as its involvement in stimulating metastatic growth of cancer cells by promoting cell survival, invasion, and angiogenesis.5,13-17 However, there have been only a few studies on periostin in relation to allergy, despite the common features of extensive tissue remodeling seen in both heart failure after myocardial infarction and allergic inflammation of airways. One previous study focused on the role of periostin in allergic eosinophilic inflammatory disease (eosinophilic esophagitis) involving extensive tissue remodeling, extending the known involvement of periostin to allergic esophagitis and proposing a novel role for periostin in the direct regulation of eosinophil accumulation in T helper type 2 (Th2)-associated mucosal inflammation of both mice and humans.18 The present study examined the expression of periostin in airway remodeling in a murine model of AR and evaluated the roles of periostin in rhinitis symptom manifestation, the inflammatory process, and airway remodeling associated with AR using periostin knockout mice.
Go to : 
MATERIALS AND METHODS
Mice and allergic rhinitis model
Mice
B6;129-Postntm1Jmol/J mice (targeted mutation of periostin) were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). Male C57BL/6J mice (4 weeks of age; the control strain for the B6;129-Postntm1Jmol/J mice recommended by the company) were obtained from Koatech Laboratory Animal, Inc. (Pyeongtaek, Korea). Homozygotes were used for experiments among the offspring generated from mating female heterozygotes and male homozygotes of the B6;129-Postntm1Jmol/J mice, according to the manufacturer's directions. All mice were housed in an air-conditioned room maintained at 22-24℃ and 50%-60% humidity. All of the animal experiments were approved by the Animal Research Ethics Board of Gyeongsang National University.
Immunization of mice and nasal challenge
Ovalbumin (OVA; Sigma, St. Louis, MO, USA)-induced airway inflammation was performed in mice as described previously.19 Male mice (C57BL/6J or B6;129-Postntm1Jmol/J) at 4 weeks of age were sensitized by intraperitoneal injection of 25 µg OVA/2 mg aluminum hydroxide gel on days 0 and 5, followed by daily intranasal challenge (from day 12 to day 19) with 3% OVA diluted in 40 µL phosphate buffered saline (PBS). Prolonged inflammation was induced by subsequent nasal exposure to 3% OVA three times a week after day 19 until the mice were sacrificed after 1 or 3 months. The B6;129-Postntm1Jmol/J strain mice were called the KO group, and the C57BL/6J strain mice were called the OVA group. Data were obtained from five mice per group. The control mice (C57BL/6J) received saline injections and saline intranasal challenges instead of the OVA solution and were called the PBS group (Fig. 1).
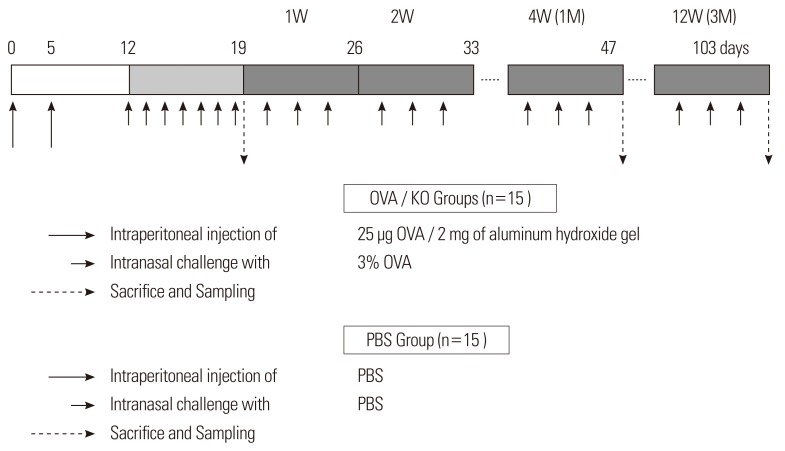
Evaluation of allergen-induced nasal symptoms
Nasal symptoms were recorded by counting the number of sneezes and nasal scratching motions (nasal rubbing) that occurred for 10 minutes after the last allergen challenge. Examinations were performed at day 19 and at months 1 and 3. The observers were blinded to the behaviors.
Preparation for histology
The experimental mice were sacrificed and decapitated after being anesthetized by an intraperitoneal injection of Zoletil® (tiletamine/zolazepam; Virbac, Carros Cedex, France). The heads were fixed in 2% paraformaldehyde and decalcified in 5% nitric acid for 5 days at 4℃. Then the heads were embedded in paraffin. The paraffin-embedded nasal cavity was transversely sectioned from the second palatal ridge to the first upper molar tooth at a thickness of 4 µm. Then the specimens were deparaffinized and stained with hematoxylin-eosin to evaluate general morphology.
Evaluation of the inflammatory process and remodeling
Eosinophils
Sirius red staining was used to visualize eosinophils. Eosinophils were counted in 10 different areas with a constant mucosal length on the slides under a high-power field (×400).
Subepithelial thickness
Specimens were also stained with Masson's trichrome to visualize collagen distribution in the nasal mucosa. Three coronal sections that were similar to the sinus cavity were chosen for evaluation. Subepithelial collagen thickness was quantified in 10 different areas at random using an NIS-Elements BR 3.0 system (Nikon Eclipse, Tokyo, Japan).
Type I collagen expression in the KO group
Immunohistochemical detection of type I collagen in the nasal tissue of the KO group was performed on 4.5-µm sections that were mounted on silicon-coated glass slides using a streptavidin-biotin peroxidase technique, as described previously.20
Quantitative analysis of matrix metalloproteinase 2, tissue inhibitor of metalloproteinase 1, and type I collagen by Western blotting
Nasal mucosal tissue lysates from five mice per group were extracted and subjected to Western blot analysis. Protein (30 µg) was subjected to electrophoresis in 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred to nitrocellulose membranes. After blocking in Tris-buffered saline/5% milk, the membrane was incubated with rabbit anti-mouse matrix metalloproteinase 2 (MMP-2) antibody (Abcam, Cambridge, UK) at a concentration of 1 µg/mL, a rabbit anti-mouse tissue inhibitor of metalloproteinase 1 (TIMP-1) antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 1:200 dilution, and rabbit anti-mouse type I collagen antibody (Abcam, Cambridge, UK) at 1:5,000 dilution, followed by goat anti-rabbit IgG horse radish peroxidase (HRP)-conjugated antibody (1:5,000). Enhanced chemiluminescence was used for autoradiography.
Identification of periostin expression in nasal tissue
Immunohistochemistry
Immunohistochemical detection of periostin in nasal tissue was performed on 4.5-µm sections that were mounted on silicon-coated glass slides using a streptavidin-biotin peroxidase technique, as described previously.20
Measurement of serum periostin levels
The blood samples were treated using the method outlined above. Quantitative assessment of periostin in serum was conducted using an enzyme linked immunosorbent assay (ELISA) kit from R&D Systems Inc. (Minneapolis, MN, USA). All of the procedures were performed according to the manufacturer's instructions. The serum samples were diluted 500-fold for the assay.
Statistical analysis
The data are presented as mean±standard error of the mean (SEM). The significance of the results was calculated using nonparametric two-sided Mann-Whitney statistical tests for comparing two groups and the Kruskal-Wallis test for comparing multiple subgroups. Statistical analysis was performed using SPSS 12.0 (SPSS Inc., Chicago, IL, USA) software. A P value of <0.05 was considered significant.
Go to :
RESULTS
Identification of periostin in the AR model
Immunohistochemical staining was performed to determine the expression of periostin and its localization in the nasal tissue of AR mice. Periostin was mainly distributed in the subepithelial tissue of the nasal mucosa as well as the pseudostratified columnar epithelium in the OVA group after 3 months of allergen challenge (Fig. 2). The area in which periostin was expressed nearly overlapped with the area that was stained by Masson's trichrome. Except for periosteal or perichondrial staining, areas positively stained for periostin were not identified in the KO and PBS groups.
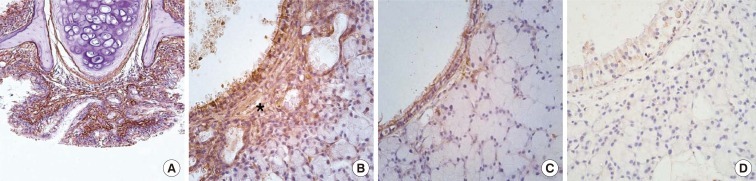 | Fig. 2Periostin immunohistochemistry. Images show the nasal respiratory mucosa after 3 months of allergen challenge. (A) OVA group, original magnification, ×20. (B) OVA group, original magnification, ×400. (C) PBS group, original magnification, ×400. (D) KO group, original magnification, ×400. Periostin was highly expressed in the subepithelial area in the OVA group (asterisk in B). However, it was less expressed in the same area of the PBS group, and no periostin expression was observed in the KO group.
PBS, phosphate buffered saline; OVA, ovalbumin; KO, knockout.
|
Serum periostin was found in the PBS and OVA groups. However, serum periostin levels were not significantly different in the PBS or OVA groups over time. None was found in the KO group, and levels were constant in all groups despite the presence of a nasal allergic response (Fig. 3).
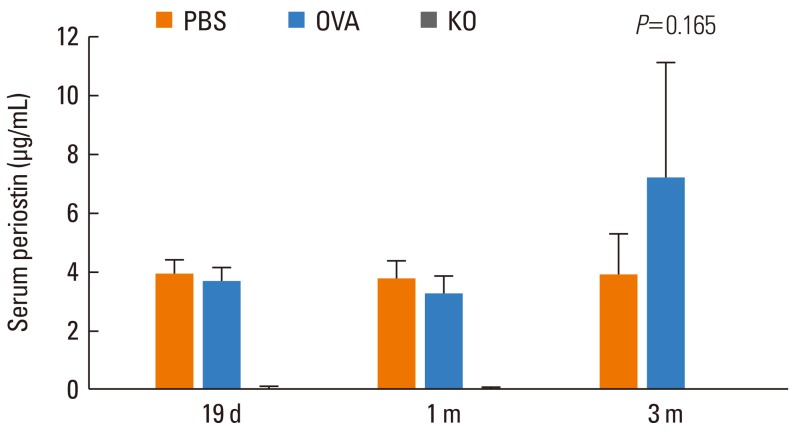 | Fig. 3Periostin levels in serum. Serum periostin levels were not significantly different in the PBS or OVA groups over time. After 19 days, 1 month, and 3 months of allergen challenge, the PBS and OVA groups showed similar serum periostin levels. Serum periostin was not found in the KO group. Data are expressed as mean±SE.
PBS, phosphate buffered saline; OVA, ovalbumin; KO, knockout.
|
Evaluation of the remodeling and inflammatory processes
Fig. 4 shows changes in the thickness of the subepithelial tissue of the nasal mucosa. It was significantly thinner in the KO group than in the OVA group (P<0.05), in which it increased gradually and peaked at 3 months (P<0.05).
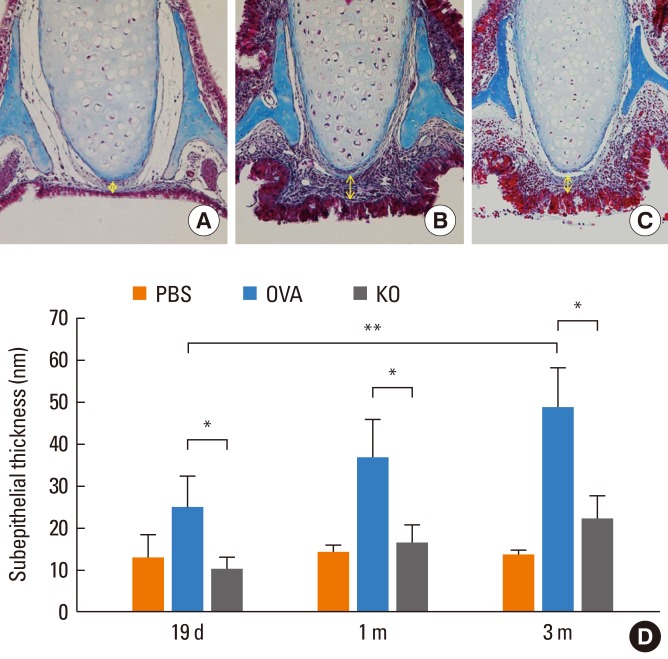 | Fig. 4Subepithelial thickness of the nasal mucosa. The subepithelial thickness of the nasal mucosa was measured (yellow arrow). (A) The nasal tissue around the septal cartilage in the PBS group after 3 months of allergen challenge (Masson's trichrome staining, original magnification, ×400). (B) The same area in the OVA group after 3 months of allergen challenge. (C) The same area in the KO group after 3 months of allergen challenge. (D) The subepithelial tissue was thinner in the KO group than in the OVA groups. *P<0.05. The subepithelial thickness increased over time in the OVA group. **P<0.05. Data are expressed as mean±SE.
PBS, phosphate buffered saline; OVA, ovalbumin; KO, knockout.
|
Collagen deposition is an important pathological feature of subepithelial fibrosis. The presence of type I collagen in the nasal tissue of the KO group was determined using immunohistochemistry (Fig. 5). Despite the deficiency of periostin, type I collagen was expressed around the subepithelial area as a matrix protein of subepithelial fibrosis. Immunoblotting was performed to evaluate the quantity of type I collagen expression in the nasal mucosa of different groups (Fig. 5). The KO group showed less type I collagen expression after 1 and 3 months of allergen challenge than the OVA group.
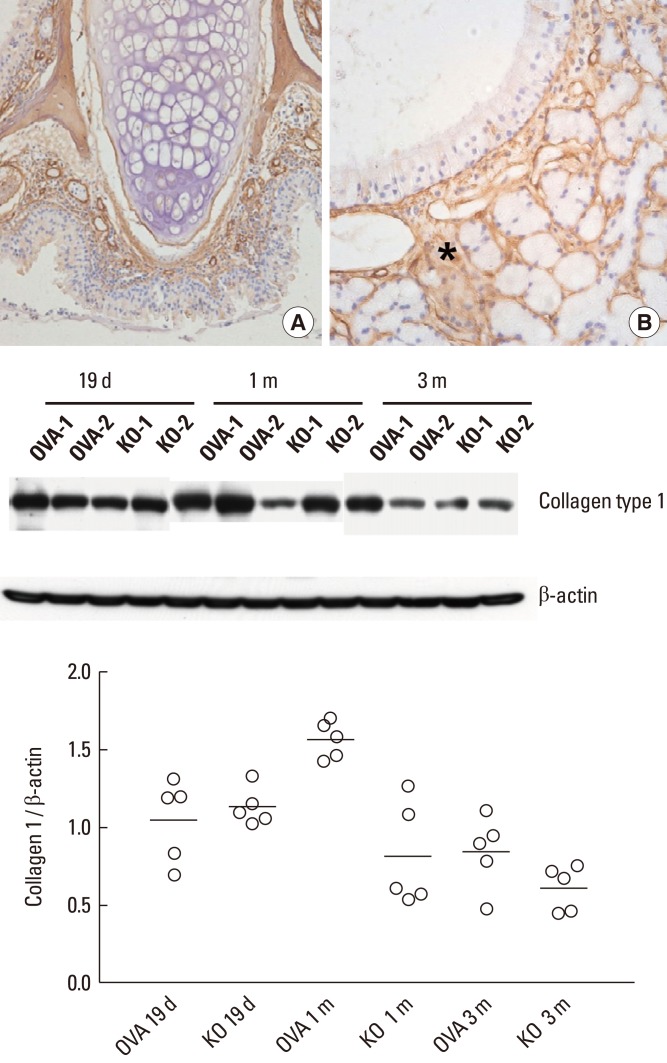 | Fig. 5Expression of type I collagen in the KO group visualized by immunohistochemistry. (A) Original magnification, ×20. (B) Original magnification, ×400. Type I collagen was identified in the subepithelial tissue (asterisk in B) of the nasal mucosa, regardless of periostin deficiency. The graph depicts the ratio of collagen type I to β-actin from nasal tissue lysates. The KO group showed a trend toward decreased type I collagen expression after 1 and 3 months of allergen challenge compared to the OVA group by immunoblotting. Two representative results of immunoblotting (n=5) over time are shown.
OVA, ovalbumin; KO, knockout.
|
To evaluate the mechanism of reduced tissue remodeling in the KO group, the expression of MMP-2 and TIMP-1, which are well-known enzymes expressed during tissue remodeling and fibrosis in the nasal mucosa, were checked by immunoblotting (Fig. 6). There were no significant differences in the expression of MMP-2 or TIMP-1 in the nasal mucosa between the KO and OVA groups over time.
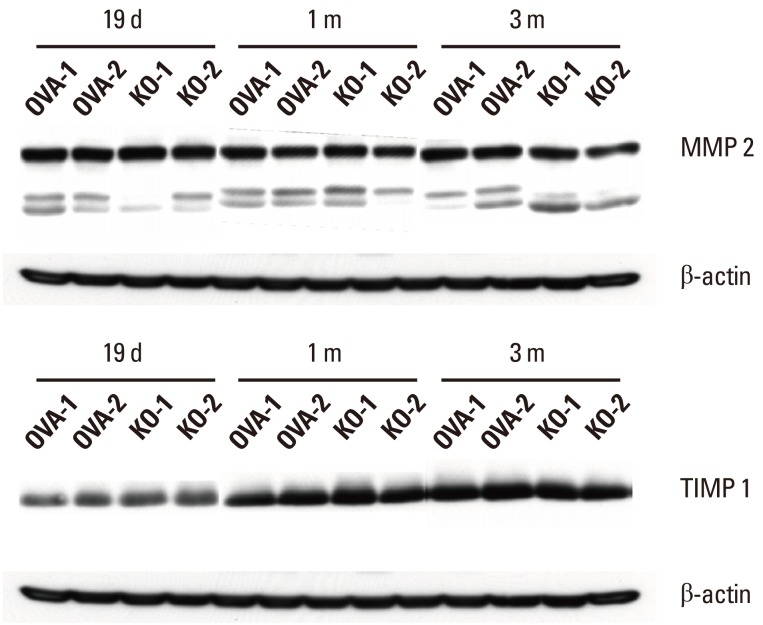 | Fig. 6MMP-2 and TIMP-1 expression in the nasal mucosa. The KO and OVA groups showed no significant differences in MMP-2 or TIMP-1 expression in the nasal mucosa by immunoblotting over time. MMP-2, matrix metalloproteinase 2; TIMP-1, tissue inhibitor of metalloproteinase. Two representative results of immunoblotting (n=5) over time are shown.
OVA, ovalbumin; KO, knockout.
|
The proinflammatory functions and the important role of eosinophils in chronic allergic diseases have been clearly elucidated.21 To determine whether periostin deficiency affects the tissue infiltration of eosinophils, these cells were counted in the histology slides after staining with Sirius red dye. The number of eosinophils in the nasal mucosa was significantly lower in the KO group than in the OVA group (P<0.05; Fig. 7). The OVA and KO groups showed significantly increased numbers of eosinophils in the nasal mucosa compared to the PBS group, which showed few eosinophils in the nasal mucosa (P<0.05). In the OVA group, the eosinophil count increased after 3 months of allergen challenge when compared to the previous time points (P<0.05).
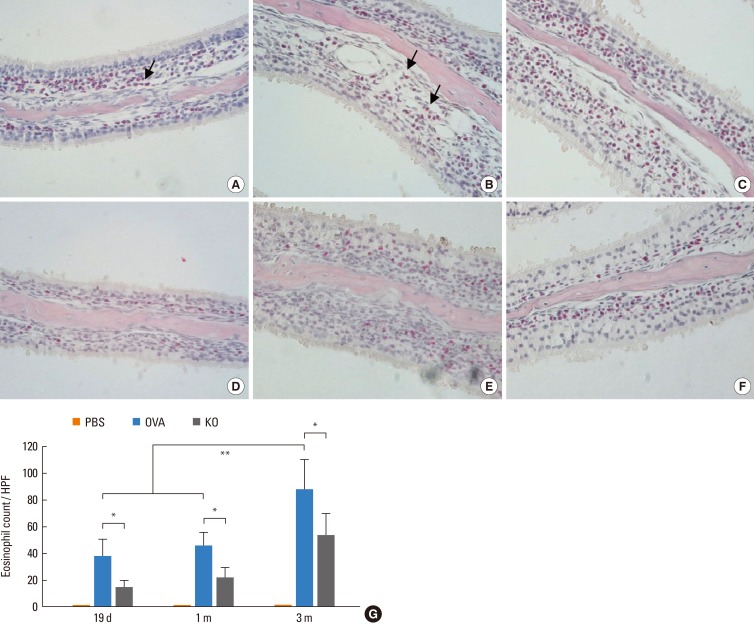 | Fig. 7Eosinophil counts in the nasal mucosa. The nasal tissue around conchal cartilage in the OVA group after (A) 19 days, (B) 1 month, and (C) 3 months are shown. Eosinophils are reddish (arrow). The nasal tissue of the KO group after (D) 19 days, (E) 1 month, and (F) 3 months are also shown (Luna staining, original magnification, ×400). (G) The number of eosinophils in the nasal mucosa was significantly lower in the KO group than in the OVA groups. *P<0.05. In the OVA group, the number of eosinophils was higher at 3 months of allergen challenge than at the other time points. **P<0.05. Data are expressed as mean±SE.
HPF, high power field.
|
Manifestation of allergen-induced nasal symptoms
Sneezing and nose rubbing, predominant features in this model after allergen challenge, were used as symptom parameters for AR. Fig. 8 shows the changes in allergen-induced nasal symptoms. The number of sneezes was lower in the KO group than in the OVA groups after 1 and 3 months of allergen challenge (P<0.05). The number of rubbings was lower in the KO group than in the OVA groups after 3 months of allergen challenge (P<0.05). The average numbers of sneezes and rubbings were higher in the OVA group than the PBS group at all recorded time points (P<0.05), and the numbers gradually increased over time. The average number of sneezes seemed higher in the PBS group than the KO group, and rubbings showed the opposite pattern, although this was statistically nonsignificant. The allergen-induced nasal symptoms were maintained during the prolonged allergen challenge.
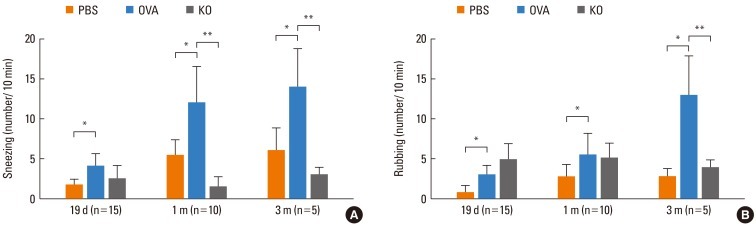 | Fig. 8Allergen-induced nasal symptoms. The numbers of (A) sneezes and (B) rubbings for 10 min immediately after the allergen challenge were counted. The OVA group had a higher symptom score than the PBS group at all time points, *P<0.05; The KO group had a lower sneeze score than the OVA group at 1 and 3 months of allergen challenge. The KO group had a lower rubbing score than the OVA group at 3 months of allergen challenge, **P<0.05. Data are expressed as mean±SE.
|
Go to :
DISCUSSION
Remodeling occurs in the nasal mucosa of AR patients,22,23 although it appears that remodeling in the nose is less extensive than in the lungs.4 One of the aims of this study was to establish a model of acute AR as well as a model of prolonged allergen-induced nasal airway inflammation to mimic the tissue remodeling that is observed in chronic AR. We focused on subepithelial fibrosis of the nasal mucosa, which increased after prolonged nasal mucosa allergen exposure in the OVA group. The pseudothickening of the reticular basement membrane caused by collagen and fibrous protein deposition occurred as a result of remodeling both in the bronchial and nasal mucosa, although the extent of pseudothickening was different between the two mucosal tissues.2
Subepithelial fibrosis is the most prominent finding in patients with eosinophilia and severe disease.24 It is also thought to change the mechanical properties of the airway wall, alter the folding behavior of the airway mucosa, heighten bronchial reactivity or narrow the airway, and reduce airflow.25,26 Multiple cytokines, growth factors, and adhesion molecules, including IL-13 and TGF-β, have been implicated in the pathophysiology of subepithelial fibrosis in asthma.27,28 In this study, the subepithelial tissue in the nasal mucosa was significantly thinner in the KO group than in the OVA groups. These results suggest that periostin deficiency may decrease subepithelial fibrosis. Three possible mechanisms for this are proposed. First, periostin itself is the main extracellular matrix protein expressed particularly in the regeneration process. Using this chronic AR model, periostin expression was identified in the subepithelial space of the nasal mucosa. Periostin was expressed during the early period of the AR model, and its expression increased with the chronicity of allergen exposure. Serum periostin levels were not significantly different in the PBS or OVA groups over time, and they were not distinctly increased after a 3-month challenge with the allergen. This suggests that the expression of periostin in this AR model might be limited to the nasal tissue. Periostin is a novel tissue component that is involved in subepithelial fibrosis in bronchial asthma6 and is also expressed in esophageal biopsies of eosinophilic esophagitis patients.18 The periostin expression in the chronic AR model used in this study is consistent with results seen in previously studied allergic diseases, and this is the first description of periostin expression in a chronic AR model. Therefore, periostin deficiency in the KO group may be the reason for the decreased subepithelial fibrosis in these mice. Second, periostin deficiency could affect epithelial mesenchymal interactions for remodeling. A previous study of human bronchial epithelial cells and fibroblasts demonstrated that periostin secreted by airway epithelial cells has autocrine effects, including the activation of TGF-β and up-regulation of type I collagen, as well as paracrine effects, including a TGF-β-mediated increase in type I collagen production in fibroblasts.29 Periostin also up-regulates MMP-2 and MMP-9 in an autocrine manner to activate TGF-β, leading to up-regulation of collagen in airway epithelial cells and fibroblasts. Changes in MMP-2 and TIMP-1 were not identified in the periostin KO group in this study. Further studies are needed to determine the effect of periostin deficiency on the epithelial-mesenchymal unit. Eosinophilic inflammation may be the third mechanism. Recent reports have supported a potential role for eosinophils in the development of airway remodeling.30-32 Increased eosinophils in the bronchial mucosa of severe asthmatics have been associated with basement membrane thickening,33 and eosinophils are capable of secreting an array of profibrotic mediators.34,35 In a study using eosinophil-deficient mice, reduced subepithelial fibrosis was independent of TGF-β expression.1 Reduced eosinophil infiltration in the KO group may be another cause of the decreased subepithelial fibrosis. Fibrosis still occurs in the periostin KO group with other extracellular matrix proteins, like type I collagen, and periostin-null mice show aberrant type I collagen fibrillogenesis in the skin and poor integrity of the periodontal ligament in response to mechanical stress.35 In the present study, the KO group showed decreased type I collagen expression after 1 and 3 months of allergen challenge compared to the OVA group. These findings suggest that decreased subepithelial fibrosis in periostin deficiency may not be attributed to the absence of type I collagen and that decreased subepithelial fibrosis might be related to decreased collagen production.
Periostin was originally identified as a gene expressed in a mouse osteoblastic cell line36; however, its pathophysiological roles remain poorly understood. It has been suggested that periostin plays a role in the development of bone, tooth, heart, and myoseptum.8,37,38 Periostin is involved in the pathogenesis of heart failure and carcinogenesis.13,39 Recently, there have been some reports about the effects of periostin in allergic inflammation. In a study of eosinophilic esophagitis, periostin directly regulated eosinophil accumulation in Th2-associated mucosal inflammation in both mice and humans.18 Periostin derived from fibroblasts or epithelial cells has been considered a mediator of inflammation and fibrosis in allergic diseases.6,18,30 However, few studies have demonstrated a pathophysiological role of periostin in nasal tissues or AR, which are highlighted by the findings in the present study.
The role of periostin in the inflammatory process of AR was evaluated in our experiments by observing the effect of periostin deficiency on a chronic AR model using periostin knockout mice. AR symptoms and eosinophil counts in nasal tissue were lower in knockout mice challenged by the allergen. These results suggest that periostin may affect or regulate the allergic response, and extend the findings of recent studies showing that eosinophil accumulation in the lung and esophagus decrease in allergen-challenged periostin-deficient mice compared to wild-type mice.18,40 It has been supposed that eotaxin-3 in the epithelium attracts eosinophils and that periostin increases the adhesion of eosinophils from the vessel to the lamina propria.18 The decrease in symptoms after 1 month of allergen challenge in the KO group might be due to reduced eosinophilic inflammation and non-specific nasal hyperreactivity. These results suggest that periostin deficiency might reduce AR symptoms in the chronic AR model.
One of the limitations of this study in explaining the effect of periostin on chronic AR is the relationship of AR to inflammatory mediators. In a previous study on periostin deficiency and allergic airway inflammation, the production of Th2 cytokines IL-4, IL-5, and IL-13 in bronchoalveolar lavage (BAL) fluid, splenocytes, and mediastinal lymph node cultures increased following stimulation with anti-CD3 or OVA from cultures of periostin-deficient cells, compared to wild-type cells.41 This study suggested that periostin deficiency could enhance Th2 inflammatory mediators in allergic airway inflammation.
The mechanisms of AR have been clarified by performing nasal challenge with allergen or proinflammatory mediators as well as by measuring cells and mediators released during the early and late phases of allergic reactions.3 However, the priming effect of the nasal mucosa is important, because a single challenge does not perfectly mimic ongoing allergic reactions induced by repeated allergen exposure. In intermittent and persistent AR, the same cells and mediators are important, but non-specific nasal hyperreactivity develops.3 Based on symptom scores, it is conceivable that periostin deficiency in this chronic AR model may reduce inflammation and remodeling.
In conclusion, periostin was expressed in the nasal tissue in a chronic murine model of AR. Periostin deficiency affected the remodeling of the nasal tissue with reduced subepithelial fibrosis. Eosinophilic inflammation in the nasal tissue was decreased in periostin-deficient mice. However, more research remains to be done on the molecular mechanism of remodeling without periostin and on acute Th2 cell-associated inflammation without periostin before this protein can be used to treat AR in clinical practice.
Go to :
 XML Download
XML Download