

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Probiotics are non-pathogenic microorganisms that confer a health benefit on the host1 by improving the balance of the intestinal microflora and possibly by augmenting host defenses. Several studies have characterized the ability of probiotic strains to alter cytokine production in the gut and associated lymphoid tissue, demonstrating immunomodulatory effects on some allergic diseases.2-4 Successful probiotic treatments in animal models of asthma have recently been described,5,6 and we have reported that oral Lactobacillus rhamnosus (Lcr35) treatment prior to allergic sensitization can attenuate airway inflammation and hyperreactivity in a mouse model of allergic airway inflammation.7 However, the beneficial effects of probiotics on the development of allergic asthma and allergic rhinitis in human clinical trials remains controversial.8,9 Despite conflicting results, it is generally thought that the immunomodulatory effect of probiotics on allergic diseases is a promising area of research.10,11
A number of studies have suggested that the immunosuppressive cytokines interleukin-10 (IL-10) and transforming growth factor (TGF)-β along with regulatory T (Treg) cells are the main mechanisms by which probiotics suppress allergic inflammation.2,5,12 Nevertheless, there have been few mechanistic studies of probiotic activity in allergic disease. In the present study, we aimed to confirm the involvement of Treg cells in the protective effect of Lcr35 in a mouse model of allergic asthma.
MATERIALS AND METHODS
Mice
Female BALB/c mice weighing 20-25 g (4 weeks old) were purchased from Orient Bio (Orient Bio Inc., Seongnam, Korea) and treated in accordance with the guidelines of the Institutional Animal Care and Use Committee (IACUC) at Asan Medical Center and Ulsan University College of Medicine.
Probiotic preparation
The Lcr35 used in this study was obtained from the Lyocentre® laboratory (Aurillac, France) and was cultured in sterile Lactobacillus MRS broth (Difco, Sparks, MD, USA) at 37℃ for 24 hours. After cultivation, cells were collected by centrifugation at 5,000×g for 10 minutes, washed twice with phosphate buffered saline (PBS), and lyophilized. The lyophilized powder was reconstituted with sterile saline before use.
Establishment of allergic asthma and probiotic treatment
Six-week-old female BALB/c mice (N=7 per group) were sensitized by intraperitoneal (i.p.) administration of an ovalbumin (OVA; 10 µg, grade V; Sigma Chemical Co., St. Louis, MO, USA) and alum (2.25 mg; Imject®, Pierce, Rockford, IL, USA) mixture. One week after the first sensitization, the mixture was administered a second time. Seven days later, the mice inhaled 1% OVA via an ultrasonic sprayer (Nescosonic UN-511; Alfresa, Osaka, Japan) for 30 minutes daily for three successive days (OVA challenge). The mice received Lcr35 (1×109 colony-forming units/600 µL/mouse/day) orally from one week before primary sensitization to the endpoint of the study. Negative controls received only saline instead of OVA at both sensitizations and airway challenge. Positive controls received nothing more after OVA sensitization.
Clinical evaluations in vivo
Airway hyperresponsiveness (AHR) in response to inhaled methacholine (MeCh; Sigma Chemical Co.), administered 24 hours after OVA challenge, was measured in conscious, unrestrained mice using a barometric whole-body plethysmograph (Buxco; EMKA Technologies, Paris, France). Briefly, mice were placed in a whole-body chamber, and basal readings were obtained for 3 minutes and averaged. Aerosolized saline followed by 5-50 mg/mL MeCh were inhaled for 3 minutes after each MeCh inhalation.
Treg cells were depleted using anti-CD25 monoclonal antibody (mAb). Briefly, mice received 250 µg of rat anti-mouse CD25 mAb (clone PC61; eBioscience, San Diego, CA, USA) i.p. in 400 µL of normal saline one day before 1% OVA challenge. Control mice were injected with 250 µg of rat IgG1 (Sigma Chemical Co.).
Bronchoalveolar lavage (BAL) fluid analysis
After measurement of AHR, the mice were anesthetized by i.p. administration of ketamine-xylazine, and the trachea was immediately exposed. The airways were lavaged through a tracheal cannula, two times with 1-mL aliquots of pyrogen-free saline warmed to 37℃. The recovered lavage fluid was pooled, and the cells were collected by centrifugation (5,000 rpm, 4℃, 5 minutes) and resuspended in 100 mL of cold PBS. The cells were stained with trypan blue to determine viability, and total nucleated cells were counted using a hemocytometer.
For differential BAL cell counts, cytospin preparations were made and stained with Diff-Quik (Sysmex, Takatsukadai, Japan). After the samples were coded, all cytospin preparations were evaluated by one observer using an oil immersion microscope (magnification, ×1,000). At least 200 cells were counted per preparation, and the absolute number of each cell type was calculated.
Serum Ig analysis
Serum was obtained from blood taken during exsanguination of the mice after airway measurement, and 100 µL (1/10 dilution in carbonate-bicarbonate buffer) were added to each well of a 96-well plate. An IgE-specific enzyme linked immunosorbent assay (ELISA) was used to quantitate total IgE in the serum, using matching antibody pairs (eBioscience) according to the manufacturer's instructions. For the ELISA, 96-well plates were first coated overnight with rat anti-mouse IgE (10 µL in 100 µL of PBS; PharMingen, San Diego, CA, USA), rat anti-mouse IgG1 (20 µg in 100 µL of PBS; PharMingen), or rat anti-mouse IgG2a (20 µg in 100 µL of PBS; PharMingen). The remaining binding sites were blocked, and the plates were incubated with 100 µL of diluted serum (1:5 for IgE, 1:10 for IgG1 or IgG2a). After the plate was washed, each of the following was sequentially added, incubated, and removed by washing: OVA (1 µg/100 µL), peroxidase-labeled rabbit anti-OVA Ig (240 ng/100 µL, PharMingen), and 3,3,5,5-tetramethylbenzidine solution (Sigma Chemical Co.). The optical density was measured at 450 nm, and the Ig level was determined relative to that of a reference pool of serum from OVA-sensitized BALB/c mice (assigned a value of 100 experimental units/mL). Determinations were performed in duplicate.
Cytokine assays
Commercial preparations of paired antibodies and protein standards for measurements of mouse IL-4, IL-5, IL-13, and IFN-γ (eBioscience) in sera were used to develop ELISAs according to the manufacturer's instructions. Determinations were performed in duplicate.
Lymphocyte proliferation assay
After BAL, the mouse spleen was resected. Mouse splenocytes were separated on a Histopaque (Sigma Chemical Co.) gradient, and the collected cells were washed with PBS. RBCs were lysed by gently mixing the cells with 3.6 mL of 0.24% NaCl for 20 seconds, followed by the quick addition of 0.3 mL of 8.7% NaCl and further dilution with PBS. The pellet was suspended in Iscove's Modified Dulbecco's Medium (IMDM), and stored overnight at 4℃. The next morning, the cells were centrifuged at 4℃, suspended in cold PBS, stained with trypan blue, and counted using a hemocytometer.
Splenic T cells were cultured in IMDM supplemented with 25 mM HEPES, 10% (v/v) heat inactivated fetal bovine serum (FBS), 60 mg/L (100 U/mL) penicillin, 100 mg/L streptomycin, and 0.29 g/L L-glutamine. Splenic T cells were adjusted to 1×105 cells/200 µL/well, transferred to 96-well plates, and incubated at 37℃ in a humidified 5% CO2 incubator for 72 hours. The cells were stimulated with OVA treatment (100 µg/mL) for 72 hours. At 12 hours before the end of the incubation, 1 µCi of [3H]-thymidine was added to each well. The cells were harvested onto a glass microfiber filter (Simport, Beloeil, Canada), and radioactivity was measured in a liquid scintillation counter. The incorporation during the last 12 hours of culture (counts per minute) was used as an index of proliferation. All cultures were tested in triplicate.
Flow cytometry
Mouse Treg cells were collected from the spleen and analyzed for CD4+CD25+Foxp3+ expression using a mouse Treg cell staining kit containing FITC-labeled anti-CD4, APC-labeled anti-CD25, and PE-labeled anti-Foxp3 (eBioscience) according to the manufacturer's instructions. Briefly, prepared cells (1×106) were washed by centrifugation with cold PBS, resuspended in 1 mL of fixation/permeabilization solution, and incubated in the dark at 4℃ for 30-60 minutes. The cells were washed once with 2 mL of permeabilization buffer, collected by centrifugation, resuspended in 20 mL of blocking agent with 2% (2 mL) normal rat serum in permeabilization buffer, and incubated at 4℃ for 15 minutes. Next, 20 mL of fluorochrome-conjugated antibody or isotype control in permeabilization buffer were added, followed by incubation in the dark at 4℃ for 30 minutes. Finally, the cells were washed with 2 mL of permeabilization buffer, resuspended in flow cytometry buffer (PBS with 2% FBS), and analyzed by flow cytometry using a FACSCalibur with CellQuest software (BD Biosciences, Mountain View, CA, USA). Determinations were performed in duplicate.
Lung histopathology
For the histological evaluation of lung tissue, the left lung of each mouse was embedded in paraffin, sectioned to a thickness of 5 µm, and stained with hematoxylin and eosin (H&E) to assess eosinophilic infiltration. Inflammation was scored by two independent, blinded investigators. The degree of peribronchial and perivascular inflammation was evaluated on a subjective scale of 0-3, as described elsewhere.13 Cellular infiltration in five randomly selected fields was assessed under a Zeiss Axiophot microscope (magnification, ×100; Carl Zeiss, Inc., Thornwood, NY, USA).
Statistical analysis
We analyzed the association between groups and positive controls using the Mann-Whitney test. Data are expressed as means±standard error. All statistical analyses were performed using SPSS ver. 18.0 for Windows (SPSS Inc., Chicago, IL, USA). A P value<0.05 indicated significance.
RESULTS
Effect of Lcr35 treatment on maximal Penh and airway eosinophilia
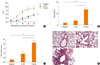
Lcr35 administration to allergic mice significantly suppressed AHR, to the level of the negative control group at the maximum MeCh dose (negative control: 4.72±2.19; Lcr35-treated: 4.75±1.56; positive control: 9.00±2.87; P<0.01) (Fig. 1A).
Lcr35-treated mice exhibited a significant suppression of BAL fluid total cell count compared with positive controls (Lcr35-treated: 4.77×105 cells/mL; positive control: 8.29×105 cells/mL; P<0.05). Oral administration of Lcr35 significantly reduced the number of eosinophils in BAL fluid compared with positive controls (Lcr35-treated: 3.79±0.29%; positive control: 13.60±3.00%; P<0.05) (Fig. 1B).
Lung inflammation after OVA inhalation was significantly reduced by oral administration of Lcr35 (Lcr35-treated: 2.16±0.55; positive control: 0.92±0.52; P<0.05) (Fig. 1C). Examination of the lung tissue of positive control mice revealed peribronchial and perivascular cellular infiltrates; Lcr35 treatment produced a marked decrease in both cellular infiltration and inflammatory changes, as determined by histopathology (Fig. 1D). Thus, Lcr35 treatment inhibited allergen-induced pulmonary inflammation, including the influx of eosinophils.
Mechanism of the effect of Lcr35 on allergic responses
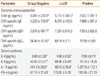
The administration of Lcr35 significantly reduced the total IgE level compared with that of the positive control, but had no significant effect on OVA-specific IgE, IgG1, or IgG2a (Table). Serum IL-4, -5, and -13 levels were significantly suppressed by orally administered Lcr35. The IFN-γ level was also decreased in Lcr35-treated mice, but the difference was not significant (Table). These results suggest that Lcr35 treatment suppresses Th2-dependent cytokines. OVA-induced cell proliferation was significantly lower in Lcr35-treated mice than in positive control mice (Fig. 2). Thus, Lcr35 effectively inhibited the OVA-specific splenic T cell response.
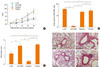
Lcr35 treatment led to a significant increase in CD4+CD25+Foxp3+ Treg cells, compared with the percentage in positive control mice (Lcr35-treated: 17.60±0.49%; positive control: 15.20±0.44%; P<0.05) (Fig. 3B). This suggests that Lcr35-induced attenuation of allergic responses in the mouse model of asthma is associated with an increased CD4+CD25+Foxp3+ Treg cell population. To confirm the involvement of CD4+CD25+Foxp3+ Treg cells, Treg cell depletion was achieved by anti-CD25 mAb treatment. Anti-CD25 mAb treatment before OVA challenge significantly increased AHR at the maximum MeCh dose, compared with AHR in Lcr35-treated mice (anti-CD25-treated: 8.16±1.63; Lcr35-treated: 3.30±0.57; P<0.05) (Fig. 4A). The isotype control rat IgG1 had no effect on AHR (data not shown). Anti-CD25 treatment also significantly decreased the population of CD4+CD25+Foxp3+ Treg cells in Lcr35-treated mice (anti-CD25 treated: 1.00±0.08%; Lcr35-treated: 15.53±0.10%; P<0.05) (Fig. 4B). These results suggest that CD4+CD25+Foxp3+ Treg cells are involved in the suppression of AHR mediated by oral administration of Lcr35.
The pulmonary inflammation score was also significantly higher in anti-CD25 mAb-treated Lcr35 mice compared with Lcr35-treated mice that did not receive anti-CD25 mAb (anti-CD25 treated: 2.28±0.19; Lcr35-treated 1.32±0.20; P<0.01) (Fig. 4C). The histopathology revealed enhanced eosinophilic inflammation in the lungs of anti-CD25-treated Lcr35 mice compared with anti-CD25 mAb-non-treated Lcr35 mice (Fig. 4D). The data indicate that administration of anti-CD25 mAb reverses the effect of oral Lcr35 treatment on lung inflammation and that Treg cells mediate the effect of Lcr35.
DISCUSSION
This study examined the effect of oral Lcr35 on allergic asthma in a mouse model. The major finding was that Lcr35 suppresses allergic parameters, including AHR, airway inflammation, and total IgE, by regulating the activity of CD4+CD25+Foxp3+ Treg cells. In addition, these effects were reversed by administration of anti-CD25 mAb.
The effect of probiotics on Th1 cytokines is controversial, as probiotics have been shown to promote, inhibit, and have no effect on Th1 cytokine production.14-16 In our study, serum levels of Th2 cytokines were reduced by oral Lcr35 treatment, whereas Th1 cytokine levels were not significantly reduced. We did not determine cytokine levels in target organs or BAL fluid; this is a limitation of this study.
To test our hypothesis, CD4+CD25+Foxp3+ Treg cell numbers in the spleen were determined. Treg cell numbers were significantly higher after Lcr35 treatment. Exacerbation readouts of asthma after Treg depletion demonstrated the importance of Treg cells in the protective effect of Lcr35. This suggests that Treg cells may be involved in the protection conferred by oral Lcr35 in the mouse model of asthma. However, other potential mechanisms such as immunosuppressive cytokines (IL-10 and TGF-β) were not examined. To our knowledge, this is the first study to identify the mechanism of action of probiotics using depletion of CD4+CD25+Foxp3+ Treg cells in a mouse model of asthma.
Some reports have suggested that the involvement of thymus-derived natural Treg cells expressing Foxp3 and CD25 in tolerance to allergens is unlikely, and the expression of Foxp3 and CD25 on Treg cells remains controversial.10 Further studies are needed to confirm the mechanism underlying the protective effect of Lcr35.
Many studies have attempted to identify the mechanism of action of probiotics in allergic diseases.2-4 Several have implicated Treg cells in the activity of probiotics,2,17 but these studies did not investigate changes in the inflammatory response after Treg cell depletion. Our study is important in that it confirms previous hypotheses that Treg cells contribute to the protective effect of probiotics.
Oral administration of Lcr35 completely blocked OVA-specific proliferation of splenic T cells in the present study. This suggests that Lcr35 regulates the systemic immune response in OVA-sensitized and -challenged mice. Oral administration of probiotics induces regulatory dendritic cells, which in turn promote the generation of CD4+Foxp3+ Treg cells in mesenteric lymph nodes.2 Dendritic cells can directly present antigens from commensal bacteria to mesenteric lymph nodes and interact with T and B cells to maintain a non-inflammatory immune response.3,4
In contrast to the massive eosinophilic infiltration in the peribronchial and perivascular areas of positive control mice, animals treated with Lcr35 showed significantly less eosinophilic inflammation. This was coincident with decreased production of the Th2 cell-derived cytokines IL-4, IL-5, and IL-13 in the Lcr35 treatment group.
There is strong evidence implicating CD4+CD25+ Treg cells in controlling allergic diseases. For example, the transfer of CD4+CD25+ Treg cells to sensitized mice reduced AHR, eosinophilic inflammation, and Th2 cytokine induction in the lung, in an IL-10-dependent manner.18,19 It is widely believed that administration of an anti-CD25 mAb results in the rapid and efficient depletion of CD4+CD25+ Treg cells, and this has been confirmed by secondary staining with a mAb directed against a different CD25 epitope.20,21 Previous studies have shown that the number of CD4+CD25+ Treg cells was significantly decreased within 3 days of anti-CD25 mAb treatment in vivo, but had recovered to normal levels by day 10.22,23 Therefore, in this study, we administered anti-CD25 mAb one day before OVA challenge. As expected, the anti-allergic effects of Lcr35 disappeared, and the CD4+CD25+Foxp3+ Treg cell population was significantly reduced. This demonstrates that the beneficial effect of Lcr35 on allergic asthma in the mice was mediated by CD4+CD25+Foxp3+ Treg cells.
In conclusion, we demonstrated that the suppression of allergic responses and immunomodulation by the probiotic Lcr35 in a mouse model of asthma was mediated by the activity of CD4+CD25+Foxp3+ Treg cells.


 XML Download
XML Download