

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Identification of a nuclear protein and an orphan receptor as IL-33 and IL-33 receptor
Interleukin-33 (IL-33), a member of the IL-1 cytokine family, is considered to be crucial for the induction of T-helper type (Th) 2 cell-dominant immune responses such as host defense against nematodes and allergic diseases.1 IL-33 was originally identified as "DVS27", a gene upregulated in vasospastic cerebral arteries after subarachnoid hemorrhage2 and as a nuclear factor, "nuclear factor from high endothelial venules (NF-HEV)", which is expressed in endothelial cell nuclei.3
IL-33 receptor was first identified as an IL-1 receptor-like molecule and termed as ST2 (the gene symbol was termed as IL1RL1) by Tominaga in 1989.4 ST2 was subsequently found to be preferentially expressed in Th2 cells and started to attract many researchers involved in allergy.5 In 2005, DVS27 was rediscovered as the 11th member in the IL-1 family of cytokines, which includes IL-1α, IL-1β, and IL-18, by computationally searching for the sequences containing β-trefoil structure seen in IL-1- and FGF-like proteins, and termed as IL1F11 or IL-33.6
IL-33 receptor and signal transduction
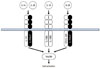
As the receptors for the other IL-1-related cytokines, the IL-33 receptor is formed heterodimeric molecules consisting of ST2 and IL-1 receptor accessory protein (IL-1RAcP; Fig. 1). IL-1RAcP is also known as a common component of receptors for IL-1α, IL-1β, IL-1F6, IL-1F8, and IL-1F9.1
The two major products of ST2 genes (IL1RL1), i.e., transmembrane form ST2 (ST2 or ST2L) and soluble form ST2 (sST2) are produced by alternative splicing under the control of two distinct promoters. ST2 is considered to be the functional component for induction of IL-33 bioactivities, while sST2 act as a decoy receptor for IL-33 like soluble IL-1Rs for IL-1.1
The signal transduction downstream of IL-33 receptor is mediated by common adapter molecules to that of the other IL-1 receptor family such as IL-1R and IL-18R. The binding of IL-33 to IL-33 receptor results in the recruitment of MyD88 to the Toll-interleukin-1 receptor domain in cytoplasmic region of ST2, leading to the induction of inflammatory mediators by activating transcription factors such as NF-κB and AP-1 through IRAK, TRAF6 and/or MAP kinases, like other IL-1 family receptor or Toll-like receptor (TLR) activation.6
THE SOURCES AND THE TARGETS OF IL-33
IL-33 as a alarmin
In contrast to the other IL-1 family cytokines except IL-1α, IL-33 is localized to the nucleus of human epithelial and endothelial cells2 and mouse bone-marrow derived cultured mast cells7 by binding to chromatin via a homeodomain (helix-turn-helix-like motif) and nuclear localization signal in amino-terminus.8 Although the pathophysiological role of IL-33 as a nuclear factor is not fully understood, IL-33 binds to the acidic pocket of dimeric histone H2A-H2B at the surface of nucleosomes, resulting in the suppression of the gene transcription at least in the in vitro reporter assay system.8
During host defense against pathogens, innate-type immune cells recognize pathogen-associated molecular patterns via TLRs, resulting in induction of inflammation. In addition, endogenous proinflammatory factors called "damage associated molecular patterns (DAMPs)" (also called "alarmin"), which are released by necrotic cells in tissue injury during trauma and/or infection, also provoke local and/or systemic inflammation by alerting acquired-type immune cells as an endogenous danger signal.9 For example, high-mobility group box 1 (HMGB1), which is originally identified as a nuclear factor as a transcriptional regulator, is released by macrophages in response to lipopolysaccharide, leading to the induction of inflammation.10 Like HMGB1, recent several lines of evidence suggest that IL-33, which also localizes in nucleus, act as a DAMP/alarmin.11
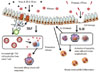
IL-33 was originally considered to be secreted by the activation of NACHT, LRR and PYD containing protein (NLRP)-mediated inflammasomes like IL-1β and IL-18 since it is cleaved from pro-IL-33 by caspase-1 in vitro.6 However, pro-IL-33 does not have a typical cleavage site seen in pro-IL-1β and -IL-18, and caspase-1 was found to proteolytically cleave pro-IL-33 at the cytokine motif, but not the intermediate region between helix-turn-helix motif and cytokine motif, resulting in the inactivation of IL-33.11-13 Like caspase-1, both caspase-3 and caspase-7 have an ability to cleave pro-IL-33 during apoptosis, in which apoptotic cells do not induce inflammation generally, and the processed IL-33 by these caspases do not have biological activities.11,12
On the other hand, biologically active pro-IL-33 can be released by necrotic cells without any processes by caspase-1, -3, -7, -8, and calpain.7,11-13 For example, pro-IL-33 can induce mouse mast cell activation to produce cytokines.13 These observations suggest that pro-IL-33 released by necrotic cells during tissue injury may have a potential role in induction of inflammation as a DAMP/alarmin.
IL-33 is an epithelial-mesenchymal cytokine manipulating inflammatory responses
Asthma is an inflammatory disease characterized by infiltration of the airway wall with a variety of immune cells and inflammatory cells such as Th2 cells, mast cells and eosinophils. However, a key component of asthma is the structural change that involves all of the elements of the airway wall associated with activation of the epithelial-mesenchymal trophic unit.14
Activated epithelial and mesenchymal cells generate a range of growth factors associated with airway remodeling and cytokines manipulating the immune response. Although IL-33 is present in the nuclei of various cell types, epithelial cells15 and endothelial cells16 are recognized as the major sources of the cytokine especially when considering the event of tissue damage.
Thus, like thymic stromal lymphopoietin (TSLP),17 IL-33 is now recognized as an epithelial-mesenchymal-derived cytokine manipulating inflammatory and/or immune responses.
Target cells of IL-33
Th2 cells
It is well established that IL-4 is a key cytokine for the differentiation of Th2 cells from naïve CD4+ T cells. ST2 is predominantly expressed on Th2 cells but not naïve T cells, Th1 cells, Th17 cells and regulatory T cells.18-20 On the other hand, ST2 is not essential for Th2 cell differentiation as shown in the study using ST2-deficient mice; ST2-deficient mice showed the normal development of Th2 cells.21,22 In support for the notion, although IL-33 cannot induce the differentiation of Th2 cells from naïve CD4+ T cells in vitro,23,24 IL-33 can enhance IL-5 and IL-13 production by in vitro-skewed Th2 cells which highly express ST2.6,25,26 Also, Kurowska-Stolarska et al.24 reported that IL-33 induces the differentiation of IL-5+IL-4- CD4+ Th cells from naïve CD4+ T cells independently of IL-4, STAT-6 and GATA-3, which are important factors for the typical Th2 cell differentiation.
Innate lymphoid cells
Lin- c-Kit+ Sca-1+ natural helper cells dwelling in the gut adipose tissue are a newly identified.29 Natural helper cells constitutively express ST2 and can produce a larger amount of IL-5 and IL-13 rather than basophils and mast cells in response to IL-33. It was shown that IL-33-mediated natural helper cell activation was important for formation of goblet cell hyperplasia during Nippostrongylus brasiliensis infection.29 Similar ST2-expressing non T/non B lymphoid cell types capable of producing IL-5 and IL-13 in response to IL-33, are subsequently identified by other investigators.30,31
Mast cells
Mast cells, which dwell in the mucosal and connective tissues, express c-Kit and high affinity IgE receptors (FcεRI), and induce IgE-mediated immune responses, are also major targets of IL-33. Mouse and human mast cells constitutively express ST2.32-35 Except IL-3 and stem cell factor (SCF, a ligand for c-kit), which are required for mast cell development at least in mouse, IL-33 is the only cytokine among 45 different cytokines which can directly provoke cytokine/chemokine (IL-1β, IL-6, IL-13, TNF, and MCP-1) secretion from mouse bone-marrow derived cultured mast cells without affecting their degranulation.36,37
Like a murine counterpart, IL-33 can induce cytokine and chemokine production, prolong survival and promote cell-adhesion in human cord blood stem cell-derived cultured mast cells.34,35 In addition, IL-33 can augment IgE-mediated cytokine production and degranulation by mouse bone-marrow derived cultured mast cells and human cord blood stem cell-derived cultured mast cells.34-36,38
Basophils
Another highly FcεRI-expressing cell type, basophils, which circulate in the peripheral blood and are potential primary sources of IL-4,39,40 are also considered as the major target of IL-33. In comparison with Th2 cells and mast cells, human and mouse basophils constitutively express ST2 at the relatively low level on their cell surface.23,26,41,42 On the other hand, the expression of ST2 on the cell surface of basophils is promoted by stimulation with IL-3.26
Like the effect of IL-33 on Th2 cells and mast cells, IL-33 alone can induce the production of cytokine including Th2-type cytokines and chemokines by basophils and promote cell-adhesion and CD11b expression in basophils in human or mice.26,27,41,42 IL-33 does not induce degranulation by basophils directly, while IL-33 synergistically enhances IgE-mediated degranulation by human basophils.26,41
In addition, IL-33 augments immune responses of basophils in human or mice; eotaxin-mediated migration,41 cytokine secretion in the presence of IL-3, which is a growth factor for basophils like mast cells,23,26,27,41-43 and prolongs survival in the presence of IL-3 or GM-CSF.41-43 These observations suggest that IL-33 is a potential activator for basophils by enhancing cytokine and chemokine secretion, recruitment and adhesion.
Eosinophils
Peripheral blood eosinophils, compared to neutrophils, are preferentially recruited into the tissue at the site of inflammation in patients with certain IgE-mediated allergic disorders such as asthma. Although ST2 expression was barely detectable on cell surface of peripheral blood eosinophils in human, ST2 mRNA and intracellular ST2 protein were detectable in them.26,44,45 IL-33 can induce the production of superoxide and IL-8 directly, and enhance IL-3, IL-5 or GM-CSF-mediated IL-8 production by human eosinophils.26,44
Like mast cells and basophils, IL-33 enhances adhesion of eosinophils by promoting CD11b expression and survival independently of IL-4, IL-5, and GM-CSF.45 Unlike basophils, IL-33 did not influence eotaxin-mediated migration of eosinophils.45 These observations strongly suggest that IL-33 may contribute to the pathogenesis of certain allergic disorders accompanied by marked accumulation of eosinophils.
Dendritic cells
IL-33 promotes the development of dendritic cells (DCs) from bone marrow cells.46 It has been shown that DCs derived by the cultivation of murine bone marrow cells in the presence of GM-CSF and IL-4 (that is, bone marrow-derived DCs; BMDCs) express ST2.47 IL-33 enhances the production of IL-6, but not IL-12, by BMDCs and augments the expression of MHC class II and CD86, but not CD80, CD40 and OX40 ligand (OX40L), on the cell surface of BMDCs.47
When naive CD4+ T cells were co-cultured with BMDCs in the presence of IL-33 for 6 to 10 days, IL-5 and IL-13, but not IL-4 and IFN-γ, were detected in the culture supernatant even without TCR engagements. Since the secreting cytokine profiles (IL-5 and IL-13, but not IL-4, production) in the settings (BMDCs + naïve CD4+ T cells + IL-33, no antigens) are similar to those by IL-5-positive IL-4-negative atypical Th2 cell population or innate lymphoid cell types, IL-33 may enhance the induction of these cell types from naïve CD4+ T cells.
Like IL-33, IL-25 and TSLP are known to be epithelial/mesenchymal cytokines inducing Th2-type cytokine-mediated immune responses.48 Contrast to IL-33, TSLP-activated DCs promotes IL-4-producing Th2 cell differentiation from naïve CD4+ T cells in the presence of TCR engagements through OX40L-OX40 interaction at least in part.49,50 IL-25 can enhance TSLP-stimulated DC-mediated Th2 cell expansion.51
Unlike IL-33, both TSLP and IL-25 can induce the differentiation of IL-4-producing Th2 cells from naïve CD4+ T cells after TCR engagements dependently of IL-4-IL-4Rα-STAT6 pathway.52,53 Therefore, these observations suggest that the role of IL-33, TSLP, and IL-25 in T cells and DCs may be different in Th2-type cytokine-mediated immune responses; TSLP and IL-25 may be involved in the preferential induction of antigen-specific IL-4/IL-5/IL-13-producing Th2 cell-mediated immune responses, while IL-33 may contribute to the induction of antigen-non specific Th2 cell-mediated immune responses by inducing IL-5/IL-13-, but not IL-4-, producing atypical Th2 cells or innate lymphoid cells.
Other cell types
Regarding epithelial mesenchymal cell types, Yagami et al.54 have examined IL-33-responsive cells among primary human lung tissue cells. They found that ST2 mRNA was expressed in both endothelial and epithelial cells but not in fibroblasts or smooth muscle cells. Correspondingly, IL-33 promoted IL-8 production by both endothelial and epithelial cells but not by fibroblasts or smooth muscle cells. Transfection of ST2 small interference RNA into both endothelial and epithelial cells significantly reduced the IL-33-dependent upregulation of IL-8, suggesting that IL-33-mediated responses in these cells occur via the ST2 receptor.
While Th2 cytokines, such as IL-4, further enhanced ST2 expression and function in both endothelial and epithelial cells, Th2/eosinophil-related cytokines/chemokines were not produced by these cell types. While the IL-33-mediated production of IL-8 by epithelial cells was almost completely suppressed by corticosteroid treatment, the effect of corticosteroid treatment on the IL-33-mediated responses of endothelial cells was only partial.54
Comprehensive role of IL-33 regarding other cell types and diseases other than asthma has been shown in our previous review article.1
IL-33 AND ASTHMA
Lessons from GWAS for asthma
The completion of the Human Genome Project, the HapMap project, and technological advances55 allowed genome-wide association studies (GWAS) to more comprehensively identify the susceptibility genes for asthma. Although asthma is now recognized as a syndrome consisting of heterogeneous disease entities,56 several genes are shown to be susceptible for asthma in GWAS. Recent large-scale GWAS all show the genes for IL-33 (IL33) and ST2 (IL1RL1) are susceptible for asthma onset.57-61 It should be noted that IL33 and IL1RL1 are located in different chromosomes, and that only these two genes are consistently listed as asthma-susceptible genes in these literatures. Interestingly, susceptible genes for atopy (IgE production) were entirely different from those for asthma including IL33 and IL1RL1.58
Thus, IL-33 now attracts much attention from all doctors and investigators who are involved in asthma research.
Role of IL-33/ST2 in asthma
The levels of soluble ST2 proteins and IL-33 mRNA/proteins are increased in sera and tissues from patients with asthma.27,62-65 Also, intraperitoneal or intranasal administration of IL-33 in mice leads to the induction of inflammation accompanied by eosinophils in mucosa of lung and intestine through the IL-13 and STAT6-dependent pathway.6,23 The levels of soluble ST2 protein and IL-33 mRNA are increased in sera and/or lungs in a murine asthma model of airway inflammation induced by ovalbumin (OVA).66,67
However, the role of ST2 and IL-33 in the induction of OVA-induced airway inflammation in mice is controversial. Especially, apparent discrepancy is often found between the studies using ST2-deficient mice and the studies using mice treated with anti-ST2 and ST2-Fc fusion proteins.1
Respiratory function, eosinophilic airway inflammation and the levels of serum total IgG1 and IgE were normally observed in 129×B6 mixed and BALB/c background-ST2-deficient mice sensitized twice with OVA emulsified with alum.21,24,68 Several investigators reported the effect of anti-ST2 mAb (clone 3E10) on OVA-induced airway inflammation in BALB/c mice (twice sensitization model with OVA/alum). Airway inflammation induced by OVA was attenuated in BALB/c mice treated with the 3E10 anti-ST2 mAb.69,70 Likewise, Th2 responses during OVA-induced airway inflammation were reduced in mice treated with anti-IL-33 polyclonal Ab.71
Adoptive transfer with DO11.10 Th2 cells, which express OVA-specific T cell receptors, into mice results in Th2 cytokine-dependent eosinophilic airway inflammation after intranasal OVA challenge.72 The airway inflammation was exacerbated when BALB/c wild-type mice or BALB/c-Rag-1-deficient mice injected with ST2-deficient DO11.10 Th2 cells were challenged with OVA in comparison with those mice injected with ST2-sufficient DO11.10 Th2 cells.68 It suggests that IL-33 signals on Th2 cells may have a regulatory function in OVA-induced airway inflammation. Contrast with the study using ST2-deficient DO11.10 Th2 cells,68 administration of the 3E10 anti-ST2 mAb or soluble ST2-Fc fusion protein in mice injected with DO11.10 Th2 cells showed the attenuated airway function and inflammation after OVA challenge.19,70
The reason for this discrepancy still remains unclear. However, it will be clarified by examining as to whether the expression of IL1RAcP, which is a signal amplifier and forms receptor complex not only with ST2 but also with IL-1 receptor (IL1R), is excessive in various immune cell types from ST2-deficient mice (Fig. 1).
Lessons from IL-33 gene deficient mice
Since the possibility of excessive IL-1 signaling in ST2 deficient mice cannot be ruled out, generation of IL-33 deficient mice has long been expected. Oboki et al.73 have only recently generated the IL-33 deficient mice. These mice normally develop, suggesting that IL-33 does not play a crucial role as a nuclear factor in physiological development.
During airway inflammation induced by twice OVA with alum sensitization, IL-33-/- mice showed attenuated eosinophil influx into the bronchoalveolar lavage (BAL) fluid and pulmonary inflammation. In contrast, IL-4 and IL-5 levels in the BAL fluid and serum OVA-specific IgE production were only slightly (i.e., not significantly) reduced in IL-33-/- mice after the last OVA challenge. IL-33-deficiency also significantly diminished inflammatory cell influx into the BAL fluid during airway inflammation induced by an extract derived house dust mites (HDM).73
Furthermore, inhalation of papain, a cystein protease allergen having strong homology with HDM allergen, Der 1,74 which can induce airway inflammation even in T/B cell-deficient Rag-2-/- mice, induced strong airway eosinophilia even without sensitization process. Most importantly, papain-induced airway inflammation is profoundly abolished in IL-33-/- mice as well as IL-4-/-IL-13-/- mice. Therefore, the papain-induced innate airway inflammation is IL-33-dependent, and that IL-4 and/or IL-13 derived from innate inflammatory cells but not T cells are important for the event.73
Taken together, IL-33 is important for inducing antigen-dependent Th2-associated local airway inflammation. However, unlike TSLP17 and IL-1, IL-33 is mostly dispensable for antigen-specific Th2 cell differentiation and antigen-specific IgE production (Fig. 2), although the actual asthma pathogenesis cannot be compartmentalized; i.e., IL-33 is capable of stimulating TSLP production.75
Oboki et al.73 also found that IL-33 is involved in the development of dextran-induced colitis accompanied by T cell-independent epithelial cell damage, but not in streptozocin-induced diabetes or Con A-induced hepatitis characterized by T cell-mediated apoptotic tissue destruction. In addition, IL-33 failed to play a substantial role in induction of T cell mediated contact and delayed-type hypersensitivity and autoimmune diseases. Of note, unnatural up-regulation of acquired immunity often seen in ST2-deficient mice is not seen in IL-33-deficient mice.73
CONCLUSION
Since recent large-scale GWAS all show the genes for IL-33 and its receptor are susceptible for asthma, IL-33 now attracts much attention from all investigators involved in asthma research. IL-33 is important for protease-mediated innate airway inflammation and in the late-phase inflammatory responses in the lung observed after IgE-mediated reaction without affecting acquisition of antigen-specific memory T cells. Thus, IL-33 provides a unique therapeutic way for asthma, i.e., ameliorating innate airway inflammation.
 XML Download
XML Download