

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Asthma is a common heterogeneous respiratory disease characterized by obstruction and symptoms that are caused by airway inflammation and remodeling.1 Inflammation and remodeling may contribute to airway hyperresponsiveness (AHR) and chronic airway obstruction.2 Current therapies targeting airway inflammation relieve and prevent symptoms in the majority of patients, although some patients experience persistent symptoms and a progressive decline in lung function, leading to the development of fixed airway obstruction and excessive airway narrowing.3,4 Patients with difficult-to-control asthma commonly take doses of inhaled therapy higher than standard doses and/or oral corticosteroid therapy.3,5 From a practical standpoint, patients whose asthma is not controlled by high-dose inhaled corticosteroid therapy are considered to have difficult-to-control/therapy-resistant asthma.3,5 Although a very small proportion of the patients with asthma have difficult-to-control/therapy-resistant asthma, these patients consume disproportional amounts of medical resources in terms of both time and money.6 What underlies this decline in lung function is unclear, but it may reflect chronic progressive airway wall remodeling. The pathological features of airway remodeling include goblet cell and mucosal gland hyperplasia, the deposition of extracellular matrix in the submucosa, smooth muscle cell hypertrophy/hyperplasia, and hyperplasia of fibroblasts/myofibroblasts.3,5 The increase in airway wall thickness, collagen content, and airway smooth muscle mass have been associated, although inconsistently, with asthma severity.7-9 The possibility that remodeling can contribute significantly to fixed airway obstruction is supported by the persistence of the latter in some patients with severe asthma under maximal treatment that eliminates airway inflammation.10 The physiological consequences of these changes remain uncertain, partially because these changes are not fully reversed by current asthma therapy.11,12 Therefore, developing a strategy to reverse airway remodeling is important so as to prevent the progressive decline of lung function in patients with asthma that persists despite long-term conventional treatments.
In recent years, interest the stromal cell system has increased; this includes marrow-derived stromal cells that support hematopoiesis, mesenchymal stem cells and their progeny, and connective tissue cells, such as osteocytes, chondrocytes, tenocytes, adipocytes, and smooth muscle cells.13 Bone marrow-derived mesenchymal stem cells (BMDMSCs) are a group of plastic adherent CD45-negative CD44H-positive cells that are capable of differentiating into a variety of cell types depending on culture conditions, including endothelial, epithelial, and neuronal cells, and adipocytes.13 When BMDMSCs are infused into mice, they can be found in the liver, muscles, heart, intestines, and lungs, with phenotypic characteristics of the cells in the organ where they reside.14 In the lungs, BMDMSCs have been detected as types I and II alveolar epithelial cells, endothelial cells, fibroblasts, and bronchial epithelial cells,15 which raises the exciting possibility that BMDMSCs may have therapeutic potential.
After venous injection, BMDMSCs must pass through the lungs. Previous studies suggest higher engraftment of BMDMSCs into the lungs compared with other organs.16,17 Previously, we reported that BMDMSCs engrafted at low levels when administered to mice challenged with bleomycin, but significantly reduced the extent of inflammation and fibrosis in the lung,13 as Ortiz et al.18 demonstrated. This suggested a therapeutic strategy using BMDMSCs in patients with severe/difficult-to-control asthma. To our knowledge, however, no trial has examined BMDMSC transfer in experimental asthma. Therefore, this study assessed the effects of BMDMSCs on airway inflammation and remodeling in a toluene diisocyanate (TDI)-induced asthma mouse model. TDI-induced asthma results in chronic airways inflammation, which increases epithelial cell damage and airway remodeling.19
MATERIALS AND METHODS
Isolation, purification, and maintenance of BMDMSCs
First, BMDMSCs were isolated from the bone marrow of Sprague-Dawley (SD) rats (Charles River Technology, Waltham, MA, USA) without contaminating hematopoietic cells, as described previously.20 Briefly, bone marrow cells were collected by flushing the femurs, tibias, and iliac crests of 6-week-old rats with phosphate-buffered saline (PBS) supplemented with 2% fetal bovine serum (FBS; GIBCO, Paisley, UK). The collected cells were cultured and maintained in T75 flasks containing Dulbecco's modified Eagle's medium and 10% FBS, supplemented with 100 IU/mL penicillin and 100 g/mL streptomycin (GIBCO). Half of the culture medium was changed at day 3 to remove non-adherent cells. Subsequently, the medium was replaced entirely each week. The cells were grown for 2-3 weeks until almost confluent. Then, the adherent cells were detached with 0.25% trypsin-EDTA. Subsequent passage and cell seeding was performed at a density of 1×106 cells. BMDMSC differentiation was analyzed using flow cytometry, as described previously.13
Intravenous infusion of BMDMSCs in a TDI-induced asthma model
Specific-pathogen-free, 6-week-old female BALB/c mice (Charles River Technology) were given 3% TDI dissolved in a mixture of ethyl acetate and olive oil (1:4) intranasally once daily for 5 consecutive days. Four days after sensitization, the mice were challenged with 1% TDI dissolved in a mixture of ethyl acetate and olive oil by ultrasonic nebulization for 1 hour over 3 consecutive days, according to a published method (Fig. 1).13,19 As a control group, sham mice were sensitized and challenged with only a mixture of ethyl acetate/olive oil. SD rat BMDMSCs (1×105 cells) were transferred to the TDI-induced asthma mice intravenously via tail-vein injection 1 day before the TDI challenge. The mice were killed on day 5 after BMDMSC transfer. The Institutional Animal Care and Use Committee of Soonchunhyang University approved the study.
Measuring AHR to methacholine and analyzing BAL cells
Airway parameters were measured 4 days after the BMDMSC transfer into TDI-sensitized mice. Mice were placed in a barometric plethysmographic chamber (All Medicus, Seoul, Korea), and baseline readings were taken for 3 minutes. The enhanced pause (Penh) was calculated according to the manufacturer's protocol [i.e., (expiratory time/relaxation time - 1)×(peak expiatory flow/peak inspiratory flow)]. Penh is a dimensionless parameter that represents a function of the proportion of maximal expiratory to maximal inspiratory box pressure signals and a function of the timing of expiration.21,22 Results are expressed as the percentage increase in Penh following challenge with each concentration of methacholine (5, 20, and 50 mg/mL). Twenty-four hours after measuring the airway parameters, the mice were killed with pentobarbital sodium (65 mg/kg, i.p.), and PBS was infused slowly into the lungs and withdrawn via a cannula inserted into the trachea to obtain bronchoalveolar lavage (BAL) fluid. The cells in the BAL fluid were counted, placed on slides by cytocentrifugation, and stained using Diff-Quick (Scientific Products, Gibbstown, NJ, USA). Then, the BAL fluid was centrifuged, and the supernatant was kept at -70℃.
Preparing lung tissue and histology and immunohistochemistry analysis
Trachea and lung tissues were removed from the mice. Then, 4% paraformaldehyde fixative solution was infused into the lungs via the trachea. The specimens were dehydrated and embedded in paraffin. For histological examination, 4-µm sections of embedded tissue were cut on a rotary microtome, placed on glass slides, deparaffinized, and stained sequentially with hematoxylin and eosin (H&E) and periodic acid-Schiff (PAS). The severity of peribronchial inflammation was graded semiquantitatively as previously described: 0, normal; 1, a few cells; 2, a ring of inflammatory cells one cell deep; 3, a ring of inflammatory cells more than four cells deep.19,23 We counted PAS-positive epithelial cells and all epithelial cells using software (Nikon DXM 1200; Nikon, Tokyo, Japan; and Image-Pro Plus 4.01; Media Cybernetics, Bethesda, MD, USA).
For immunohistochemistry of smooth muscle actin (SMA) and proliferating cell nuclear antigen (PCNA), lung tissue on slides was treated with 0.3% H2O2 for 30 minutes to block endogenous peroxidase and then incubated at 4℃ overnight with anti-mouse SMA goat polyclonal antibody (1:200 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and anti-mouse PCNA rabbit polyclonal antibody (1:200 dilution; Santa Cruz Biotechnology). After the slides were incubated with an avidin-biotin complex kit (ABC; Vector Laboratories, Burlingame, CA, USA), color was developed with 3,3' diaminobenzidine (DAB; Zymed Laboratories, San Francisco, CA, USA).
Masson trichrome staining of lung tissue and measuring collagen in lung protein extracts
Lung tissues were fixed in Bouin's solution, washed in tap water (5 minutes at room temperature), and then stained for 10 minutes with Weigert's iron hematoxylin. After washing in tap water, the slides were stained with a mixture of 1% acid fuchsin and 1% Biebrich scarlet in distilled water for 2 minutes and then treated with 2.5% phosphomolybdic-phosphotungstic acid for 10 minutes. Finally, the sections were stained with aniline blue for 1 minute, treated with 1% acetic acid for 1 minute, and then dehydrated in an ethanol/water series followed by five changes in absolute ethanol, cleared in xylene, and mounted in balsam. The collagen assay was performed according to the Sircol collagen assay kit manual (Biocolor, Carrickfergus, Northern Ireland, UK). Briefly, 100 µL lung tissue protein extract was mixed with 1 mL of Sircol dye for 30 minutes and centrifuged at 10,000 rpm for 5 minutes to drop the formed collagen-dye complex. After decanting the suspension, droplets were dissolved in 1 mL Sircol alkali reagent and vortexed. The acquired solution (100 µL) was read at 540 nm, as previously described.13
Statistical analysis
Data are expressed as the mean±standard error of the mean. SPSS version 10.0 (SPSS, Chicago, IL, USA) was used. Continuous data were compared using the Kruskal-Wallis test. If differences were found to be significant, the Mann-Whitney U-test was applied to compare differences between two samples. Differences were considered significant when P<0.05.
RESULTS
Effect of BMDMSCs on airway inflammation in a TDI-induced asthma model
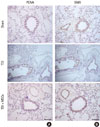
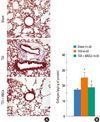
The BMDMSCs used in this study expressed CD44H and CD45 on the surface based on flow cytometry analysis, as described previously.13 TDI-sensitized/challenged mice had a significantly more cells and fractions of inflammatory cells in BAL fluid compared with sham mice (P<0.05; Fig. 2). BMDMSC transfer significantly reduced the TDI-induced increase in the number of cells, neutrophils, eosinophils, and macrophages in BAL fluid to the level of that in sham-treated mice (P<0.05; Fig. 2). On histologic examination, TDI-sensitized/challenged mice had a heavy infiltration of inflammatory cells and exudative changes in the peribronchial layers and intraluminal areas of the bronchi. BMDMSC transfer significantly reduced the cellular infiltration induced by TDI-sensitization/challenge (Fig. 3A). The semiquantitative value of the inflammatory index from H&E-stained images was significantly attenuated by the infusion of BMDMSCs in the TDI-sensitized/challenged mice (Fig. 3B).
Effect of BMDMSCs on goblet cell numbers, collagen deposition, and SMA and PCNA protein expression in the airways
The percentage of PAS-positive goblet cells (Fig. 3C) increased significantly in the TDI-sensitized/challenged mice, and the BMDMSC infusion significantly reduced the number of goblet cells to the sham level (P<0.0001). PCNA protein expression (Fig. 4A) markedly increased in the peribronchial muscle layer and in the epithelial cells in the bronchi of TDI-sensitized/challenged mice, and this was effectively reduced by BMDMSC transfer. TDI-sensitized/challenged mice also strongly expressed SMA protein in the bronchial and intraluminal exudations of the intrapulmonary bronchi (Fig. 4B).
To quantify collagen deposition in the lungs, Masson trichrome staining and the Sircol collagen assay were used. As shown in Fig. 5A, collagen deposition increased in the lungs from TDI-sensitized/challenged mice, and the infusion of BMDMSCs into mice reduced the TDI-induced collagen deposition. The result concurred with the finding that BMDMSC-infused mice had significantly lower levels of measurable collagen than did the TDI-sensitized/challenged group (P<0.005; Fig. 5B), reaching the normal level of the sham group.
Effect of BMDMSC transfer on the airways of a TDI-induced asthma model
The Penh values increased significantly in TDI-sensitized/challenged mice compared with the sham mice at 20 and 50 mg/mL methacholine (P<0.005 and P<0.01, respectively; Fig. 6). The BMDMSC infusion reduced the increased Penh values of the TDI-sensitized/challenged mice at 50 mg/mL methacholine to the level of sham controls (P<0.01; Fig. 6).
DISCUSSION
This study evaluated the effects of BMDMSCs on both inflammation and remodeling in a TDI-induced mouse asthma model. TDI is a low-molecular-weight compound used widely in the production of polyurethane foams, automobile paint, varnishes, and related products. TDI-induced asthma may reflect a subtype of severe asthma.24 Approximately 5-10% of workers in these settings develop occupational asthma with chronic airways inflammation. Therefore, we used a TDI-induced asthma mouse model as a subtype of severe asthma.
In this study, the BMDMSC infusion inhibited the TDI-induced increase in the numbers of macrophages, eosinophils, and neutrophils (Fig. 2), which suggests that BMDMSCs inhibit inflammatory cell infiltration non-selectively. Mice exposed to TDI developed features of airway remodeling, including thickening of the peribronchial smooth muscle layer, subepithelial collagen deposition, and increased airway mucus production.12 In our study, SMA and PCNA stains were used to measure myofibroblast and smooth muscle thickening and epithelial cell damage and regeneration associated with remodeling. Notably, the BMDMSC infusion effectively inhibited the increased SMA-positive muscle layer, and PCNA-positive epithelial cells induced by TDI-sensitization/challenge (Fig. 4), suggesting that BMDMSCs effectively inhibit airway remodeling in asthma. In addition, the semiquantitative value of the inflammatory index and the percentage of goblet cells induced by TDI-sensitization/challenge were significantly reduced after BMDMSC transfer, suggesting that the infusion of BMDMSCs inhibits not only airway inflammation, but also mucus metaplasia. The BMDMSC infusion significantly reduced the levels of measurable collagen compared with the TDI-sensitized/challenged group, indicating that the infusion of BMDMSCs can protect remodeling in TDI-sensitized/challenged mice.
In the lungs, many studies have reported that either whole bone marrow or BMDMSCs limited experimental lung injury and fibrosis,13,18,25 but the idea that stem cells simply supply a reservoir of new lung parenchymal cells and therefore hasten repair is inadequate to explain the effect.13 Although one can demonstrate the persistence of donor stem cells in the lungs and evident differentiation into lung cell phenotypes, the number of engrafted cells is too small to explain the protective effect sufficiently.25 Similarly, infusions of whole bone marrow protect endotoxin-induced lung injury at a level disproportionate to donor cell number found in the lungs.26 The anti-inflammatory effect of BMDMSCs may be an important contributor to the inhibitory effect of stem cell administration.27
The anti-fibrotic effect of BMDMSCs has been demonstrated in bleomycin-induced pulmonary fibrosis.13 In the inhibitory mechanism of BMDMSCs, both endogenous and exogenous stem cells contribute to the repair process.25 Homing of stem cells to injured lung is at least partly a result of the production of humoral mediators by injured, but not by normal, lung that are chemotactic for stem cells.25 We did not evaluate the mechanism of BMDMSC effectiveness, and further studies are needed to clarify whether the effect of BMDMSCs is due to BMDMSCs or to humoral mediator release.
In summary, BMDMSC transfer effectively inhibited inflammation and remodeling in a TDI-induced asthma mouse model, suggesting that BMDMSCs, which possess anti-inflammatory and anti-remodeling properties, could be useful for the treatment of inflammation and remodeling in severe asthma. To date, there has been no report on the effects of BMDMSCs on inflammation and remodeling in experimental asthma. This study is the first to establish the therapeutic effects of BMDMSCs in a TDI-induced asthma model.




 XML Download
XML Download