

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The main role of the immune system is the ability to distinguish self from non-self while still responding to and neutralizing pathogens. The physiopathology of immune tolerance-related diseases, such as allergy, asthma or autoimmune diseases is complex and influenced by several factors. These include genetic susceptibility, the nature of the antigen that initiates the disease (antigen dose, time of exposure, route of exposure, and its structural characteristics) and possible co-exposure with innate immune response stimulating substances, such as infections and flora bacteria.1
Since allergy is a predominantly Th2 type of immune disorder, one of the essential ways to overcome the deviated immune response has been allergen-specific immunotherapy (SIT) that involves repeated administration of the sensitizing allergens by subcutaneous injection or mucosal route. The induction of peripheral tolerance towards the responsible allergen is the main target in allergen-SIT. After successful immunotherapy, allergen-specific T regulatory cells (Treg) are generated and they suppress proliferative and cytokine responses against allergens.2 In addition, antibody class-switching occurs in B cells that secrete allergen-specific IgG4 instead of IgE which has blocking capacity inhibiting binding between allergen and IgE on mast cells and basophils. This review focuses on mechanism of allergen-SIT and discusses the current approaches in clinical and research perspective of immune tolerance induction in allergic disorders.
IMMUNE RESPONSE TO ALLERGENS
Loss of tolerance to certain allergens such as, aeroallergens, foods and insect venom, leads to induction of type I hypersensitivity reactions. The type of immune response influenced by several factors including genetic susceptibility, the nature of antigen which initiates the disease (antigen dose, time of exposure, route of exposure, and structural characteristics), and challenge with infections and bacteria.3 Under these complex stimulus, navie T cells activated by professional antigen-presenting cells (APC) and differentiate into Th1, Th2, Th17 or Th9 cells. For atopic disease, the Th2 arm of immune response is responsible for immunopathology and clinical scene. Once Th2 response is established, the mechanism of allergic disease is further divided into two main phases: first sensitization, and development of memory and later followed by effector phase and tissue injury. In the sensitization phase, allergen-specific CD4+Th2 cells produce IL-4 and IL-13, which induce B cell class-switch into the antibody isotypes of ε immunoglobulin heavy chain and the production of allergen-specific IgE antibody. Later, allergen-specific IgE, binds to high affinity receptor for IgE (FcεRI receptors) on the surface membrane of mast cells and basophils. These series of activations lead to the sensitization of the patients to a specific allergen. Re-exposure to the sensitized allergen leads to the aggregation of receptor-bound IgE molecules and results in the activation and mediator release that lead to the development of clinical symptoms of type I hypersitivity reactions.4,5
Immediate reactions are followed by late phase reactions with the activation of T cells by continuous presence of allergens. Once allergen-specific CD4+Th2 cells are activated, they produce IL-4, IL-5, IL-9, and IL-13, which play crucial role in the maintenance of high allergen-specific IgE levels, stimulate eosinophil progenitors in the bone marrow, induce inflammatory cell influx into inflamed tissue, and induce production of mucus and smooth muscle contraction.3 These events require T cell activation and peripheral T cell tolerance prevents formation of atopic immunopathology in healthy subjects. A continuous interaction with migrating T cells and resident tissue cells takes place and other subsets such as Th1 cells, Th9 cells, Th17 cells, and Th22 cells also play important roles.6-8
ROLE OF TH17 AND TH22 CELLS ON ALLERGIC DISORDERS
Th17 cells represent a newly discovered subset of T lymphocytes and are involved in the pathogenesis of several immune-mediated disorders. IL-17, IL-6, TNF-α, and IL-22 are signature cytokines of Th17 cells and play important roles on tissue pathology in autoimmune disorders as well as allergic disease.9 Recent studies showed the importance of allergen-specific Th17 cells in humans.10 IL-17 is essentially important for the recruitment of neutrophils and expressed in bronchial biopsies, bronchoalveolar lavage fluid and sputum of patients of asthma.11 IL-17A and IL-17F are negative regulators of antigen driven Th2 response.12,13 It has also been demonstrated that anti-IL-17 reduces neutrophilic infiltration in experimental murine asthma models.14 Furthermore, IL-17 increases eosinophilic infiltration and recruitment/survival of airway macrophages.12 Patients with allergic rhinitis revealed high frequency of IL-17-producing T cells.15 Taken together, IL-17 contribute to the differentiation and activation of allergen-specific Th2 cells, influx of eosinophils to target organs, and serum IgE production that provide important clues in the role of Th2 driven allergic response.16
Similar to Th17 cells, Th22 cells were rarely found in PBMCs, while they were clearly detected in T cell population isolated from skin of patients with psoriasis, atopic eczema and allergic contact dermatitis.17 Furthermore, IL-22 synergizes with IL-17 in the induction of proinflammatory cytokines in human bronchial epithelial cells18 and colonic myofibroblasts.19
DENDRITIC CELLS AS KEY PLAYERS IN ALLERGIC INFLAMMATION
Dendritic cells (DCs) play a crucial role for detecting innate pathogens and triggering adaptive immune response. Within the subtypes of DCs, plasmacytoid DCs (pDCs) take attention by their surface phenotype, tissue localization, cytokine secretion, and antigen-presentation function,20 and keep important role in the initiation and regulation of immune response and immune tolerance.21 The maturation period influences the role of DCs in the tolerance formation. Especially, semi-mature DC induce tolerance in the immune system, whereas matured DCs mediate expression of interferon (IFN)-γ producing T cells.22 Taken together, distrubiton of DC subsets and their maturation stage seems to be essential in tolerance induction to certain antigens.
Murine oral mucosal dendritic cells (mDCs) express CD11b+/CD11c-/+ at the mucosal/submucosal junction zone, whereas langerhans cells epress CD207+ within the mucosal epithelium and pDCs express B220+/120G8+ in the submucosal region. In a study on sublingual application of OVA, a total clearence of OVA within 15-30 minutes by CD11b+/CD11c-/+ mDCs was shown.23 It seems however that these murine studies are not fully relevant to human in vivo stimulation. In humans, oral mucosal Langerhans cells (oLCs) represent the predominant DC population, however pDCs are virtually absent in oral mucosa. oLCs constitutively express high affinity receptor for IgE, which is absent in classical epidermal Langerhans cells. FcεRI expression is seen during early differentiation period of Langerhans cells as well as other DCs and it possesses a pro-tolerogenic character. Studies clearly demonstrate that oLCs of atopic individuals show increased expression of FcεRI that cooperates with IgE.24 This strategic location of oLCs at suprabasal epithelium layer and increased expression of FcεRI may facilitate binding and processing of allergens in sublingual immunotheraphy (SLIT) period.
ROLE OF REGULATORY T CELLS IN IMMUNE TOLERANCE TO ALLERGENS
The importance and functions of Treg cells to induce tolerance have been explicitly studied during the last 15 years. The major role of Treg cells in immune tolerance was clarified in murine studies directly or adoptive transfer of Treg cells. They prevent or cure numerous T-cell-mediated disease models, including, asthmatic lung inflammation, autoimmune diseases and allograft rejection, by achieving immune tolerance to responsible allergens, self antigens or alloantigens.25 On the other hand, chronic absence or imperfect function of Treg cells may lead a series of immune dysfunction disease, such as hyper IgE syndrome, hyper eosinophilia and autoimmunity in humans, which is normal with appropriate function of Treg cells.26 For an easy understanding, Treg cells may be divided into two main subgroups; The naturally occurring forkhead box P3 (Foxp3)+CD4+ CD25+ regulatory T cells (will be referred as CD4+CD25+Treg cells)27, which develop in the thymus and are present in birth, and the other is inducible Treg cells, which is generated in the periphery under various tolerogenic conditions. Especially, the IL-10-producing T regulatory type 1 (Tr1) cells have been shown to play a key role in allergen tolerance, and can be induced by allergen-SIT in humans.28-31 With recent studies, it is well established that Foxp3 acts as master switch transcription factor for Treg cell development and function.32 Foxp3 mutations in the mice leads spontaneous development of allergic airway inflammation, hyper IgE syndrome, eosinophilia as well as autoimmune disease.26 Mutations in Foxp3 gene in humans leads formation of X-linked immune dysregulation polyendocrinopathy enteropathy syndrome (IPEX), hyper IgE syndrome and eczema.26 A dysregulation of disease-causing effector T cells is observed in atopic dermatitis lesions, in association with an impaired CD4+CD25+FoxP3+T-cell infiltration, despite the expression of type 1 regulatory cells in the dermis.33 Apart from these, main subsets of Treg cells, several other T cells with regulatory function has been described. Among them, suppressor capacity in vitro revealed CD8+ CD28- T cells, which are able to prevent up-regulation of B7 molecules induced by Th cells on professional APCs34 and play role in oral tolerance.35 TCRαβ+CD4-CD8- double-negative Treg cells have been shown to suppress Ag-specific immune responses mediated by CD8+ and CD4+ T cells in humans and mice.36 NKreg cell also has the capacity to suppress antigen specific T cell response.37 It has recently been demonstrated that, the transforming growth factor-beta (TGF-beta) induced the expression of the Runt-related transcription factors RUNX1 and RUNX3 in CD4+ T cells.6 This induction seems to be a prerequisite for the binding of RUNX1 and RUNX3 to three putative RUNX binding sites in the FOXP3 promoter. Further, in this study it was shown that RUNX transcription factors act as a molecular link in TGF-beta-induced Foxp3 expression in inducible Treg cell differentiation and function.
Tr1 cells are dominant type of T cell subset in healthy individuals. Studies clearly show that, allergen-specific Tr1 cells are predominant in healthy individuals to prevent unwanted immune response to nonpathogenic environmental antigens such as house dust mite, birch pollen, bee venom and food antigens (hazelnut, pear) which lead to allergy.38,39 Healthy and allergic individuals denote three different allergen-specific T cell subtypes as Th1, Th2 and Tr1 in different ratios.39 The imbalance between Th2 and Tr1 cells and depending the dominant subset may conduce allergy development or recovery from allergy. Peripheral T cell tolerance to venom allergen is an appropriate model for high dose tolerance to allergens in humans. During beekeeping season repeated exposure of non-allergic healthy beekeepers to bee venom antigens denote an efficient model to apprehend mechanisms of immune tolerance to bee venom allergens.40 During the exposure to venom allergen, venom specific IL-10-secreting Tr1 cells show a switch from allergen-specific Th1 and Th2 cells. This leads to suppression of allergen-specific undesired immune response by Th1 and Th2 cells. This immunomodulator response persist as long as venom exposure continuous and returns to the initial level within 2-3 months after the end of the beekeeping season and stimulation of histamine receptor 2 on Th2 cells by histamine suppresses allergen-stimulated T cells and enhance IL-10 production as an additional immune tolerance mechanism. Supporting these findings, non-allergic beekeepers have approximately 1,000 times higher allergen-specific IgG4 versus allergen-specific IgE ratio compared to bee venom allergic individuals.41 Another tolerance model with cat allergen also showed elevated levels of allergen-specific IgG4 levels after exposure to high dose cat allergen.42
THERAPEUTIC APPROACHES THAT USE REGULATORY T-CELLS
With the knowledge of suppressive and immunomodulatory capacity of inducible or constitutive Treg cells, novel treatment strategies for T-cell mediated disease such as transplantation rejection, autoimmunity and allergy are being developed. The curative and preventive effect on disease conditions experienced by both adaptive transfer of regulatory T cells or their induction by immunomodulators in vivo are under consideration. Compared to conventional treatment strategies, their antigen-specific suppressor capacity as well as long-lasting antigen-specific regulation in vivo with a limited side effects were reported.43 Recent studies on immunomodulators that is targeting to enhance or suppress the numbers and functions of Treg cells, are rapamycin,44 co-stimulatory blockage,45 non-mitogenic anti-CD3 mAbs,46 T cell depletion47 and anti-TNF-α mAb48 (Table).49-75
Specifically expanded Treg cells can be targeted to allergen or an autoantigen expressed in the inflamed organs in murine models.76 Further, the transfer of this organ specific Treg cells can suppress an ongoing disease.77 These studies aimed for a successful therapeutical approach by targeting the Treg cell arm of immune tolerance against allergens, autoantigens or transplantation antigens. Several investigations are ongoing for adoptive transfer of Treg cells or small compounds aimed to induce Treg cells in the tissues.76 Other therapeutical approaches such as allergen-SIT, treatment with glucocorticoids, and beta-2 agonists seem to particially function by promoting the numbers and activity of IL-10-secreting Tr1-like cells.78-80
SUBCUTANEOUS OR SUBLINGUAL ROUTES OF ALLERGEN-SPECIFIC IMMUNOTHERAPY
Allergen-specific immunotherapy, repeated administration of the sensitizing allergens by subcutaneous injection or mucosal route, has been used nearly 100 years ago by Noon and Freeman to grass polen allergic patiens with grass polen extracts. Clinical and experimental studies clearly show that SLIT is relatively safe method compared to subcutaneous immunotheraphy (SCIT) for the treatment of allergic disease,81,82 however the mechanisms of SLIT are less understood and its efficacy seems to be less than SCIT.83-85 Oral mucosal tissue has a natural tolerogenic character without any acute inflamation in spite of high bacterial colonization and good wound healing without scar development. Lack of inflammatory cells around mucosal tissue and high permeability for allergens suggests a way of action for sublingual allergen immunotherapy.86 The first step of SLIT is to uptake an allergen by Langerhans cells-specialized dendritic cells24 within the oral mucosa via high affinity surface IgE receptors.87 This leads to secretion of IL-10 and induction of T cells with a regulatory phenotype in vitro.88 Although clinical trails demonstrate treatment efficacy of SCIT in allergic asthma, allergic rhinitis, and stinging insect hypersensitivity as well as aero-allergen-induced atopic dermatitis such as reduction of allergic symptoms and drug intake, there is a risk of serious adverse reactions which can be classified in two categories: local reactions, which can appear as erythema, pruritus and swelling at the injection site; and systemic reactions can range in severity from mild to very severe life-threatening anaphylaxis.89-91
MECHANISMS OF ALLERGEN-SPECIFIC IMMUNOTHERAPY
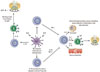
The primary purpose of allergen-SIT is the induction of peripheral T cell tolerance to allergens.28-31,39,92 Once peripheral T cell tolerance is triggered, allergen-specific Treg cells produce elaveted levels of IL-10 and TGF-β which are anti-inflammatory cytokines.29-31,38 The cytokines secreted from Treg cells mostly depend on the type of organ they dwell and the path in which they are stimulated. Experimental and clinical data revealed that Treg cells may secrete only IL-10, IL-10, and IFN-γ or IL-10 and TGF-β.25,39,93-96 Treg cells do not only suppress Th2 immune response and peripheral tolerance is achieved with multiple mechanism to overcome and suppress allergic inflammation. The other roles of Treg cells are suppression of dendritic cells and by this, enhance the generation of effector or induction of dendritic cells that support the generation of Treg cells,97-99 suppression of Th1 and Th2 cells,100 suppression of allergen-specific IgE and induction of IgG4 and/or IgA,101 suppression of mast cells, basophils and eosinophils,102 interaction with resident cells and remodelling.103,104 In SCIT both circulating30 and mucosal Tregs105 form and these Tregs may induce B cells to produce protective IgG4 antibodies106 and IgA2 antibodies.107 Proliferative response of T cell clones are also inhibited by IgG4 antibodies through prevention of IgE-faciliated allergen binding to B cells and subsequent presentation to allergen-specific T cell clones108 (Figure).
Mucosal immunotherapy to house dust mite, birch polen or food antigens leads to induction of Tr1 cells, which secrete IL-10 and TGF-β.31 Treg cells stimulated with toll-like receptors produce IL-10 and IFN-γ.94 After venom allergen-SIT, there is a induction of Tr1 cells producing only IL-10.109 Overall, it is obvious that allergen-SIT has a modulatory effect on allergen-specific T cells. In the mechanism of succesful allergen-SIT, shift in Th2 immunity to Th1 immune response is observed in peripheral blood,110,111 allergic rinitis112 and cutaneous late phase responses.110 One of the studies carried on patients with allergic rihinitis revealed that after grass pollen immunotherapy, Foxp3+ CD25+ and Foxp3+CD4+ cells numbers were found to be increased in the nasal mucosa.105 Also after this allergen-SIT, IL-10-producing Tr1 cells increased with supporting the role of Treg cells in the induction of allergen-specific tolerance in the humans.105 A very recent study showed that SIT with grass pollen extract leads to an increase in Foxp3+ cells in the sublingual epithelium.113
Treg cells and antibody response in allergen-SIT
B cell activation and antibody production is an essential event during an immune response to antigens. Mature B cells show antibody class-switching in response to antigenic stimulation and costimulatory signals.114 Isotype class-switching of B cells depends mainly on cytokines released from T helper cells. In allergic immune response, Th2 cells secrete high amount of IL-4 and IL-13, which lead to ε-chain switching of B cells and the secretion of elevated levels of IgE. Other factors affecting the isotype swithing are CD40 and/or TLR stimulation. Even though peripheral T cell tolerance is rapidly induced after SIT, whether B cells tolerance takes place is still uncertain. Clinical data revealed that normal immune response to allergen results in increased production of allergen-specific IgE together with allergen-specific IgG4. In allergen-SIT, a transient increase in serum specific IgE115 is followed by gradual decrease over months or years of treatment. In addition, allergen-specific IgG4 and IgG1 subtypes of IgG antibody increase 10 to 100-fold in serum levels with successful allergen-SIT.49 Secreted IgG4 is thought to block the allergen before its binding and cross-linking of the IgE on the surface FcεRI receptors of mast cells and basophils, and by this inhibits activation and release of mediators responsible for type I hypersensitivity. However, there is poor correlation between the amount of allergen-specific IgG and clinical protection. Consequently, it becomes important to measure the blocking activity of allergen-specific IgG4 as well as IgG1 instead of their serum levels. In a clinical study of SLIT with grass polen extract, it was shown that IgG1 and IgG4 levels as well as seasonal IgA1 and IgA2 levels are increased.113 Also in this study, there is significant inhibition IgE-faciliated allergen binding to B cells at peak season, and time dependent increase in serum inhibitory activity for IgE-FAb in SLIT-treated atopic individuals.
IL-10 secreted from regulatory cells has a sophisticated role on the immune response with downregulation of T cells and induction of allergen-specific IgG4 antibodies. IL-10 counter-regulates allergen-specific IgE vs IgG4 levels.28,101 Thus, IL-10 regulates tolerance in T cells as well as inducing the IgG4 isotype rather than IgE phenotype. Clinical studies manifest during allergen SIT to sensitized patients, although there is no significant change in specific IgE levels, there is significant increase in specific IgA, IgG1, and IgG4 levels.31 Further studies also showed the correlation between increase of IgA and TGF-β and between increase of IgG4 and IL-10 in peripheral mucosal response to allergens in healthy individuals as well as patients treated with allergen-SIT.31,40 During allergen-SIT IgE/IgG4 ratio decreases and this is associated with a change from allergen-specific Th2 to Treg cell predominance. The change in IgE/IgG4 ratio is observed almost after several weeks due to early increase in IgG4.28 This is probably due to long living IgE plasma cells in the bone marrow. Significant decrease in IgE levels occurs in years in spite of early generation of Treg cells. Overall, allergen-SIT has immunomodulatory effect on antigen-specific T cells and B cells, and leads to early remission of clinical and latephase responses with a switch towards IgG4 in allergen-specific antibody levels.
CONCLUSION
In this review, we discussed the recent developments and more established knowledge on mechanisms of allergen-SIT. Peripheral T-cell tolerance is the key immunologic mechanism in the healthy immune response to self and non-infectious, non-self antigens. Induction of peripheral T cell tolerance by Treg cells is the main event that takes place in successful allergen-SIT. It is characterized by induction of Treg cells, suppressive cytokines such as IL-10 and also non-inflammatory antibody isotypes including IgG4 and IgA are essential to overcome allergic state. Knowledge of this molecular basis is pivotal in understanding the equilibrated regulation of the immune response and unresponsiveness to immunologic agents and their possible therapeutic applications. A crucial area for future studies is the identification of drugs, cytokines, or costimulatory molecules that induce peripheral T cell tolerance to environmental allergens. Novel vaccines that shorten the duration, decrease side effects, increase efficiency for treatment as well as novel preventive approach are expected due better understanding the mechanism of immune tolerance.

 XML Download
XML Download