

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Toluene diisocyanate (TDI) is a very common cause of occupational asthma (OA) worldwide. The pathogenic mechanisms of TDI-induced asthma are more variable than those of atopic and non-atopic asthma. However, similar to non-OA, OA may result from the complicated interactions of multiple genetic and environmental factors.1,2 Genetic factors may modify TDI-induced asthma susceptibility, as seen in previous studies in which only 5%-15% of isocyanate-exposed workers developed asthma.3 The genes for glutathione S-transferase (GSTP1 and GSTM1),4 N-acetyltransferase (NAT),5 HLA classes I and II,6,7 and neurokinin 2 receptor (NK2R)8 may all confer susceptibility to or protection against TDI exposure-associated asthma. β2-adrenergic receptor gene (ADRB2) polymorphisms have been identified, which are functionally relevant and may modify disease; numerous clinical studies have described the effects of such polymorphisms on asthma and related phenotypes.9,10 The ADRB2 is a small, intronless gene that has recently been resequenced in multiple ethnic populations to determine polymorphic variability and haplotype structure.11 Two ADRB2 single nucleotide polymorphisms (SNPs) at codons 16 and 27 are frequently associated with the asthmatic phenotype; the Gly16 variant is positively associated with severe and nocturnal asthma.9,12,13 Isocyanates may act as pharmacological inhibitors of β2-adrenergic receptors in TDI-OA by affecting the ability to produce cyclic adenosine monophosphate and to maintain bronchial tone.14,15 A previous study established that β2-adrenergic receptor stimulation inhibited release of proinflammatory mediators from mast cells and influenced T-cell growth and function as well as eosinophil survival and function.16 Therefore, many lung cell types implicated in asthma pathogenesis are potentially influenced by β2-adrenergic receptor signals. Previous studies have identified pharmacoethnic differences between Korean and Caucasian populations in the ADBR2 genotype distribution and the pharmacogenetic association of responses to β2-agonists.17-19
No studies to date have evaluated ADRB2 polymorphisms as possible disease modifiers in TDI-OA. To our knowledge, this is the first investigation to evaluate the genetic association of ADRB2 polymorphisms with the TDI-OA phenotype in a Korean population.
MATERIALS AND METHODS
Subjects
Patients with confirmed TDI-OA (n=103) were included in the study. Patients were primarily diagnosed by positive responses to TDI bronchoprovocation tests. TDI-exposed asymptomatic workers (n=60) were enrolled from work environments with similar intensities of TDI exposure (spray painters, furniture polishers, and musical instrument factory workers) and comprised the asymptomatically exposed control group (AEC). A total of 263 unexposed healthy control subjects (NC) were recruited from the volunteer population. All subjects provided informed consent and the study was approved by the Institutional Review Board of Ajou Medical Center, Suwon, Korea.
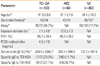
The demographic data for the three study groups are summarized in Table 1. Atopy was determined by a positive skin test to at least one common inhalant allergen. The battery of allergens included the house dust mite, tree and pollen mixtures, mugwort, ragweed pollens, and alternaria (Bencard, Worthing, West Sussex, UK). Serum samples from TDI-OA individuals were collected at initial diagnosis during the exposure period; all participants discontinued inhaled or oral steroids 4 weeks prior to the start of the study. Participants in the TDI-OA and NC groups underwent an interview, chest radiography, and skin prick test with common inhalant allergens. Pulmonary function measurements and methacholine inhalation challenge were performed in TDI-OA subjects.
Bronchial challenge test with methacholine and TDI
The methacholine bronchial challenge test was performed in TDI-OA patients as described previously.20 Briefly, aerosols were generated with a DeVilbiss 646 nebulizer connected to a DeVilbiss dosimeter driven by compressed air (DeVilbiss Co., Doylestown, PA, USA). Normal saline inhalation at 5-min intervals was followed by a series of progressively increasing doses of methacholine (0.075-25 mg/mL) until a 20% fall in forced expiratory volume in the first second (FEV1) was observed or until the maximum dosage was reached. FEV1 was measured 5 min after the initiation of each set of aerosolized methacholine inhalation. The methacholine concentration resulting in a 20% fall in FEV1 (PC20) was determined by interpolation from the dose-response curve. The TDI bronchial challenge test was performed according to the procedure described previously.20
Specific IgG and IgE antibodies to TDI-HSA conjugate determined by ELISA
The vapor-type TDI-human serum albumin (HSA) conjugate was prepared from 80% 2,4-TDI isomers based on the recently described isocyanate vapor-phase exposure system.21,22 Serum-specific IgE and IgG levels were determined by enzyme-linked immunosorbent assay (ELISA), as described previously.21 Briefly, TDI-albumin conjugate (1 µg) or mock conjugate was dissolved in normal saline and used to coat ELISA plates (Corning, New York, NY, USA) at 37℃ for 2 h, followed by overnight incubation at 4℃. Plates were washed with PBS-Tween 20 (PBS-T), and blocked with blocking buffer (350 µL, PBS containing 5% BSA and 0.05% Tween 20). Sera from patients and control subjects were diluted 1:5 for specific IgG measurements and at 1:3 for specific IgE; sera (50 µL) were incubated in TDI-albumin- and mock albumin-coated wells for 1 h at 25℃. Plates were washed four times, and alkaline phosphatase-conjugated anti-IgG antibody (1:20,000, 100 µL; Sigma, St. Louis, MO, USA) or goat anti-IgE (1:1,000; Kirkegaard and Perry Laboratories, Gaithersburg, MD, USA) and alkaline phosphatase-conjugated rabbit anti-goat IgG antibody (1:200; Kirkegaard and Perry Laboratories) was added to each well and incubated for 1 h at room temperature. Plates were washed, and substrate solution (100 µL) was added to each well (Sigma); plates were incubated for 30 min at room temperature. The absorbance at 405/450 nm was determined using an ELISA plate reader. Specific TDI binding was calculated as the difference in optical density between mock-conjugated and the TDI-albumin wells.
Identification and genotyping of single nucleotide polymorphisms
The ADRB2 gene fragment containing the promoter and exon 1 (1,483 bp) was sequenced to examine SNPs in DNA samples from subjects. DNA sequencing was performed using an ABI Prism 3100 DNA analyzer (Applied Biosystems, Foster City, CA, USA). Primers (5'-CCTAAAGTCCTGTGCACATAAC-3' and 5'-GCCCTCAGATTTGTCAATCTTC-3') were designed for amplification and sequencing. The PCR mix (30 µL) contained primers (10 pmol), dNTPs (1 µM), and Taq DNA polymerase (1 U). The amplification program included a single cycle at 94℃ for 5 min followed by 35 cycles of 94℃ for 30 s, 60℃ for 40 s, and 72℃ for 90 s, with a final extension at 72℃ for 7 min. Sequence variants were identified on chromatograms, which detected six SNPs (-47 T>C, -20 T>C, Arg16gly A>G, Gln27Glu C>G, Leu134 Leu G>A, and Arg175Arg C>A). The SNP genotypes were verified by direct sequencing of the PCR products.
Statistical analysis
The chi-square (χ2) test with one degree of freedom (df) was used to test the frequency of each SNP for significant departure from Hardy-Weinberg equilibrium. Differences in genotypic frequency between patients or exposed and control subjects were examined by the χ2 test. To evaluate the genetic associations among the TDI-OA, AEC, and NC groups, logistic regression models (codominant, dominant, and recessive) were used to analyze SNPs and haplotypes after controlling for age and gender as covariates. Subject haplotypes from ADRB2 polymorphisms were estimated using the hapassoc package in R 2.9.0,23 based on an estimation-maximization algorithm (EM). Linkage disequilibrium between loci was measured with the logarithm of odds (LOD) score. Logistic regression controlling for age and gender as covariates was applied to predict the genetic effects of ADRB2 haplotypes for IgE sensitization to TDI-HSA conjugate using the null haplotype as the reference. The P-values were corrected by false discovery rate (FDR, R2.7.1; R foundation for Statistical Computing, Vienna, Austria), which is a statistical means to correct for confounding effects of multiple genetic and clinical factors. Associations are expressed as adjusted logistic odds ratio (OR) and 95% confidence intervals (95% CI). Student's t test was used to compare continuous variables (values are expressed as means±standard deviation [SD]). Categorical variables were compared using the χ2 test or Fisher's exact test. Continuous variables without a normal distribution were log-transformed and presented on their original scale. Statistical analyses were performed with SPSS ver. 12.0.1 (SPSS Inc., Chicago, IL, USA). In all analyses, P<0.05 was taken to indicate statistical significance.
RESULTS
Demographics of the study population
The demographic data of TDI-OA patients (n=103), AEC individuals (n=60), and unexposed healthy controls (NC; n=263) are shown in Table 1. There were no significant differences in mean age, gender distribution, or serum total IgE levels between the TDI-OA and AEC groups. Exposure durations were significantly longer in the AEC group than in the TDI-OA group. Methacholine challenge testing was not performed in participants in the AEC and NC groups because these healthy individuals reported no respiratory symptoms.
Allele, genotype, and haplotype frequencies of the beta 2-adrenergic receptor gene
A genetic association study of six SNPs (-47 T>C, -20 T>C, Arg16Gly A>G, Gln27Glu C>G, Leu134Leu G>A, and Arg175Arg C>A) was performed in the three study groups (Table 2). The allele and genotype frequencies were consistent with previous studies in Korean populations.24,25 There were no significant differences in allelic, genotypic, or haplotypic frequencies of the six SNPs among the study groups. The six SNPs of ADBR2 polymorphisms were in linkage disequilibrium (LOD score >3, Figure); four haplotypes including ht1 [TTACGC], ht2 [TTGCAA], ht3 [CCGGGC], and ht4 [TTGCGC] were constructed with the EM algorithm, but both ht3 and ht4 had a frequency of less than 5%. There were no significant differences in genotype distributions of the four ADRB2 haplotypes among the three groups (Table 2).
Effects of ADBR2 polymorphisms on the clinical parameters in patients with TDI exposure
We compared the clinical parameters in TDI-exposed subjects (TDI-OA and AEC groups) according to the six ADBR2 SNPs to assess the relationship between ADBR2 polymorphisms and the TDI-OA phenotype (Table 3). There were no significant differences in asthma duration, baseline FEV1, PC20, or total serum IgE levels among the genotypes of the six SNPs (data not shown).
Among the three analysis models (codominant, dominant, and recessive models), the ADRB2, Arg16Gly, Leu134Leu, and Arg173Arg polymorphisms showed highly significant effects on the prevalence of serum specific IgE to the TDI-HSA conjugate, with the highest significance observed in the codominant model (OR, 3.09; 95% CI, 1.47-6.51 for Arg16Arg A>G, OR, 2.42; 95% CI, 1.16-5.07 for Leu134Leu G>A and OR, 2.97; 95% CI, 1.39-6.32 for Arg173Arg C>A) (Table 3). Of the six SNPs, only the Arg16Gly A>G polymorphism had a significant association with IgE sensitization to the TDI-HSA conjugate under all three modes of inheritance. This effect was significant after correction for multiple hypothesis testing (P=0.038 for the codominant model, P=0.041 for the recessive model). Homozygous ADRB2 ht1/ht1 showed a significantly higher prevalence of specific IgE to TDI-HSA conjugate compared with the null ht1 (P=0.048; OR, 15.40; 95% CI, 1.81-131.06; Table 3). However, the ADRB2 ht2 haplotype was not significantly associated with IgE sensitization to the TDI-HSA conjugate in this study. The statistical power (type I error=0.05) for evaluating the specific IgE to TDI-HSA conjugate association with genetic polymorphisms (ADRB2 Arg16Gly A>G and Arg175Arg C>A and ht1 [TTACGC] was 0.795, 0.745, and 0.858, respectively.
No significant differences were observed with serum specific IgG antibody.
Homozygotes for the ADRB2 Arg16 allele or haplotype 1 [TTACGC] tended to show lower PC20 methacholine than Gly16 homozygotes among TDI-OA patients (homozygous for Arg16, 3.1±5.3 mg/mL vs. homozygous for Gly16, 9.3±17 mg/mL; P>0.05) in terms of airway hyperresponsiveness assessed by the methacholine bronchial challenge test. However, there was no significant relation between the genotypes or haplotypes of the six ADRB2 SNPs and the degree of airway hyperresponsiveness.
DISCUSSION
We screened the genotypic and allelic frequencies of six ADBR2 SNPs, including the two most common sites, Arg16Gly and Gln27Glu, in the present study. We identified significant associations between the frequencies of the Arg16Gly and Arg173Arg SNPs and the ht1 [TTACGC] haplotype and the prevalence of specific IgE antibodies to the TDI-HSA conjugate among TDI-exposed participants. Several genetic factors have recently been identified that may alter OA risk. The present study is the first to investigate a possible association between ADBR2 polymorphisms and a specific phenotype of adult-onset OA in a Korean population. Our data suggest that the ADBR2 haplotype ht1 [TTACGC], including the Arg16 allele, contributes to the development of specific IgE sensitization to TDI among TDI-exposed Korean workers.
The β2-adrenergic receptor is a G protein-coupled receptor that mediates catecholamine actions in multiple human tissues. β2-adrenergic receptors are present on the surface of inflammatory cells and may modulate signaling pathways in these cells. β2-adrenergic receptor stimulation could induce increased IgE production by human B lymphocytes; it may also enhance production and expression of soluble, high-affinity IgE receptor (sCD23), suggesting that this receptor agonist may be involved in IgE synthesis.26 The anti-inflammatory effects of β2-agonists may inhibit the elaboration of mediators from mast cells.27 Several studies have demonstrated that altered β2-adrenergic receptor signaling due to the ADBR2 polymorphisms Arg16Gly and Gln27Glu could potentially affect airway smooth muscle contractility and inflammatory cytokine production, and therefore modulate IgE production through β2-adrenergic receptors expressed on B and T lymphocytes in vitro and in vivo.28-30 A previous study demonstrated that increased adrenoceptor downregulation was related to the ADRB2 Arg16Gly polymorphism.31 However, other studies have not identified significant associations between the Gly27Glu or Arg16Gly ADRB2 polymorphisms and total serum IgE levels in Korean populations.17,25,32 Llanes et al.33 did not identify significant associations between genetic variations of ADRB2 Arg16Gly polymorphisms and the risk of olive pollen sensitization and asthma. Conversely, Ramsay et al.34 reported that the ADBR2 Arg16Gly polymorphism (Arg16 variants) was associated with a significantly increased specific IgE level and increased wheal size in response to skin prick testing with Ascaris lumbricoides in a Venezuelan cohort, although no significant associations were identified between physician-diagnosed asthma and ADRB2 genetic polymorphisms in the same population.
It is generally accepted that TDI exposure can induce specific IgE, resulting in TDI-OA.3,21,22,35 We detected IgE with the 80% vapor-type TDI-HSA conjugate in the present study, and demonstrated a significant association with the ADBR2 Arg16Gly AA/AG genotype and ht1 [TTACGC]; these observations suggested that this genotype could serve as a risk factor for increased specific IgE sensitization in TDI-exposed workers and could ultimately contribute to the development of asthmatic symptoms. However, the mechanism by which the ADBR2 polymorphism induced IgE sensitization to inhaled TDI is still unclear. It is possible that β2-adrenergic receptors on inflammatory cells from subjects with the haplotype 1 [TTACGC] of ADBR2 polymorphisms, and particularly the ADRB2 Arg16Gly AA genotype, are more susceptible to the generation of serum specific IgE antibodies in response to inhaled TDI.
Our study had possible limitations considering the issue of multiple comparisons. The six ADRB2 SNPs evaluated in the present study had moderate levels of linkage disequilibrium and were thus not fully independent tests. In this case, if strict Bonferroni correction was applied to the multiple comparisons for a single genetic polymorphism, false negative rates usually increased. Therefore, an alternative adjustment for multiple hypothesis testing is to use the FDR method, which estimates the likelihood that our findings are false.36 Regardless of FDR adjustments, evidence of significant associations among ADRB2 Arg16Gly, Arg175Arg, and ht1 with the prevalence of specific IgE to TDI-HSA conjugate remain.
OA is a complex disease characterized by multiple genetic and environmental factors. We attempted to reduce environmental effects on the asthma phenotype in the present study by enrolling healthy, asymptomatic exposed controls. After adjusting for age, gender, and other ADRB2 SNPs, the ht1 [TTACGC] haplotype, composed of all wild genotypes of Arg16Gly, Leu134Leu, and Arg175Arg among the six ADRB2 SNPs, was significantly associated with specific IgE sensitization in TDI-exposed workers. Additional environmental factors, including inhaled aeroallergen exposure, smoking, and viral infections, must undergo further evaluation.37 Therefore, further studies are required to evaluate the associations between ADRB2 polymorphisms and TDI-OA pathogenesis and to better elucidate the complex interactions of ADRB2 polymorphisms with environmental or genetic factors that may increase TDI-OA susceptibility. Our results suggest that the ht1 haplotype, which includes all wild genotypes of Arg16Gly, Leu134Leu, and Arg175Arg, may be a risk factor for developing specific IgE sensitization to TDI.



 XML Download
XML Download