

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Despite many undeniable therapeutic successes, breast cancer remains a major health issue in both developed and developing countries. Obesity, as reflected by increased body mass index (BMI), is associated with an increased risk of more aggressive breast cancer, as well as reduced survival [1]. The association between BMI and breast cancer risk becomes particularly strong for postmenopausal women [2] and weight gain after menopause is the most important risk factor for breast cancer in postmenopausal women [34]. However, the 2007 WCRF/AICR Second Expert Report [5] followed by the Updated Breast Cancer 2010 Report [6] found limited evidence on the association between breast cancer risk and dietary components which may contribute to body fat mass. The association between dietary fat has been a topic of debate for more than 20 years, however a recent meta-analysis reported no significant association [78].
Estrogen is a known modulator of lipid and glucose metabolism. Systemic loss of estrogen in postmenopausal women is typically associated with increased abdominal fat tissue. Studies have indicated that estradiol (E2) replacement can prevent menopause induced gains in adipose tissue mass [910]. In addition, ovariectomized (OVX) rodents rapidly become obese; however estrogen administration prevents the increase in body fat [11]. Estrogen activates AMP-activated protein kinase (AMPK), promoting fat oxidation and decreased expression of sterol regulatory element-binding protein 1c (SREBP1c) in adipose tissue, muscle, and the liver [1213]. SREBP1c stimulates expression of lipogenic genes, such as fatty acid synthase (FAS) [1214]. In addition to lipid storage in adipose tissue, estrogen deprivation disturbs glucose homeostasis. For example, premenopausal women are more insulin sensitive, have insulin-associated improved glucose tolerance, and are less likely to develop insulin resistance than men [1516]. OVX rodents were reported to show dyslipidemia, impaired glucose tolerance, and impaired insulin-mediated glucose uptake in skeletal muscle [17]. In addition, estradiol replacement therapy improved glucose tolerance and insulin sensitivity and reduced lipid accumulation in the liver of leptin-deficient ob/ob mice [18]. One study suggested postmenopausal women with increased risk of hyperinsulinemia, insulin-resistant type 2 diabetes, and metabolic syndrome [19]. Association of postmenopausal weight gain with hormone-dependent breast cancer development has also been demonstrated [20]. Previous studies have reported overexpression of the insulin receptor (IR) in breast cancer cells [21]. Increased insulin binding to the IR leads to stimulation of the phosphoinositide 3-kinase (PI3K)/AKT signaling pathway, which plays an important role in breast cancer progression [22]. AKT is transported to both cytosolic and nuclear compartments, where it phosphorylates various proteins involved in the regulation of cell growth and apoptosis. Abnormal AKT activation has been observed in various types of cancer [2324]. Therefore, it can be hypothesized that defects in insulin sensitivity in postmenopausal women and an increase in fat mass may play a role in breast cancer development. However, few mechanistic studies have provided evidence to explain the association between excess energy intake in breast cancer development under conditions with different estrogen availability.
In this study, we evaluate the effects of estrogen deprivation and/or excess energy intake on mammary tumor development and growth in a rodent model. Possible molecular mechanisms of action are also suggested.
MATERIALS AND METHODS
Animals and experimental design
Three-week old C3H/HeN female mice were obtained from Central Laboratory (Seoul, Korea). Animals were housed in polycarbonate cages and maintained at a room temperature of 26 ± 1℃, with a relative humidity of 60 ± 5%, and 12 h light/12 h darkness exposure. Fresh food was provided every 2-3 days and food intake was monitored throughout the experiment. Body weight was monitored once a week.
After acclimation for one week, animals were either ovarectomized or sham-operated and then randomly assigned to different experimental groups. Group 1 consisted of shamoperated animals fed a normal diet (15% energy as fat) (SHAM+ ND, n = 14); Group 2 consisted of sham-operated animals fed a high-fat diet (45% energy as fat) (SHAM+HD, n = 8); Group 3 consisted of ovariectomized animals fed a normal diet (OVX+ND, n = 6); and Group 4 consisted of ovariectomized animals fed a high-fat diet (OVX+HD, n = 7). The composition of the experimental diet was based on AIN-93G diet (25) as shown in Table 1, and soybean oil was substituted with corn oil because isoflavones present in soybean oil may act as anticarcinogens. A mixture of lard and corn oil was used as a source of dietary fat to mimic the fatty acid composition in the human diet as used in our previous study (26). Mineral mix, vitamins, choline, and tert-butylhydroquinone were adjusted to provide an equal amount for each experimental group based on difference in daily food consumption of animals in ND and HD. At 7 weeks of age, all animals were provided with a dose of 1mg 7,12-dimethylbenz(a)anthracene (DMBA) (Sigma, MO, USA) dissolved in sesame oil. This dosage was repeated once a week for six consecutive weeks. After the last DMBA exposure, mice were subjected to regular examination for mammary tumors by palpation. Tumor size was measured using a digital caliper, and the length and width of each tumor were used in the formula [volume = length2 × width/2] to approximate volume (cm3). Mice were sacrificed at the age of 19 weeks. All mammary tumors were weighed and the size was measured. All procedures were approved by the Institutional Animal Care and Use committee of Sookmyung Women's University (SMU-IACUC-2010-0625-009).
Preparation of blood and tissue samples
At the end of the experiment, animals were sacrificed and the mammary tumor, mammary fat pad, liver, spleen, and abdominal fat pad were removed, rinsed in normal saline, and weighed. Blood was collected from the inferior vena cava into EDTA-free tubes and centrifuged at 1,550 × g for 20 minutes. All samples were stored at -80℃ until assayed.
Serum measurements
Serum insulin concentration was determined using a commercially-available enzyme-linked immunosorbent assay (ELISA) kit (Millipore, MA, USA). Serum free fatty acid (FFA) concentration was also determined using an ELISA kit (Wako, Osaka, Japan) according to the manufacturer's instructions.
Western blots
Protein expression of IR, mammalian target of rapamycin (mTOR), p-mTOR, AKT, pAKT, AMPK, and phosphorylated AMPK (pAMPK) was measured in mammary tumor tissue samples. Because only two animals in the OVX+ND group developed tumors, statistical analyses were performed in SHAM+ND, SHAM+HD, and OVX+HD animals. Tumor tissue was homogenized with a PRO-PREP™ protein extraction solution (Intron Biotechnology Inc., Gyeonggi, Korea), left on ice for 20 minutes, and centrifuged (16,600 × g, 10 min, 4℃. Protein content was determined against a standardized control, using a Bio-Rad Protein Assay kit (Bio-Rad Laboratories, Inc., CA, USA); 50 µg of protein from each sample was separated by 4-12% and 6% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to PVDF membranes (Koma Biotechnology, Seoul, Korea). The membranes were blocked with 2% skim milk (Amersham Corp., IL, USA) and incubated with specific antibodies for IR (Santa Cruz Biotech, CA, USA), pAKT (Ser473) (Cell Signaling, MA, USA), AKT (Cell Signaling, MA, USA), mTOR (Cell Signaling, MA, USA), pAMPK (Cell Signaling, MA, USA), AMPK (Cell Signaling, MA, USA), cyclin D1 (Cell Signaling, MA, USA), CDK4 (Cell Signaling, MA, USA), and β-actin (Sigma, MO, USA). The membranes were washed with PBS/Tween 20 (PBST) containing 0.1% Tween 20 (Sigma, MO, USA). Reactive bands were visualized using an enhanced chemiluminescence (ECL) system (Amersham Corp., IL, USA). Stripping was checked by re-exposure to enhanced chemiluminescence (ECL), and was detected using a LAS 3000 (Fujifilm, Tokyo, Japan). The membranes were subsequently blocked and reprobed. The intensity of the bands was quantified using a Bio-Rad GS-800 densitometer equipped with the Quantity One program (Bio-Rad Laboratories, Inc., CA, USA).
Real-time quantitative PCR
Total RNA was extracted from liver tissue using TRIzol reagent, according to the manufacturer's instructions (Invitrogen, CA, USA). Total RNA (1 µg) from liver tissue was reverse-transcribed using a cDNA Synthesis kit (PhileKorea Technology, Seoul, Korea) according to the manufacturer's instructions. First strand cDNA was generated from 1 µg RNA using both oligo (DT)18 primer mix and random hexamer primer mix. Real-time quantitative PCR was performed on a 7500 Fast Real time PCR system (Applied Biosystems, CA, USA) using a QuantiMix SYBR Kit (PhileKorea Technology, Seoul, Korea). Primers for SREBP1c, FAS, and Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were synthesized by Bioneer (Bioneer, Daejeon, Korea) (Table 2). Relative fold-changes in expression were determined using the 2-ΔΔCT (relative quantification) analysis protocol. Expression of the GAPDH housekeeping gene was used to normalize PCR reactions. Each experiment was repeated three times.
Statistical analysis
Statistical analysis was performed using the SAS package (release 9.1, SAS Institute Inc., NC, USA). Data are expressed as the mean ± SD. One-way analysis of variance (ANOVA) and Duncan's multiple test were used to determine statistical differences between the treatment groups. Interactions between two variables were examined using two-way ANOVA. P-values less than 0.05 were considered significant. Only two mice developed mammary tumors in the OVX+ND group; therefore, statistical analyses on tumor tissue were performed for animals in the SHAM+ND, SHAM+HD, and OVX+HD groups using one-way ANOVA.
RESULTS
Body and organ weights
There was no significant difference in body weights between experimental groups (Table 3). No significant difference in the weights of liver, spleen, abdominal adipose tissue, or mammary fat pad was observed. Neither diet nor OVX affected body weight and organ weights was observed between groups.
Mammary tumor development
At the end of the study period, the percentage of tumor-bearing mice in each group was 57.1% in the SHAM+ND group, 75% in the SHAM+HD group, 33.3% in the OVX+ND group, and 42.9% in the OVX+HD group (Table 4). Number of tumors of tumor bearing mice in each group was 1.63, 1.50, 1.50, and 1.67, respectively. The average mammary tumor volume was significantly higher in the OVX+HD group than in the other groups. Both diet and OVX showed significant association with tumor size, and there was no significant interaction between ovariectomy and diet.
Serum concentration of insulin and free fatty acids
Significantly higher circulating concentrations of insulin were observed in the OVX+HD group compared with the other groups (Table 5). A significant difference in FFA concentration was observed between the OVX+HD group and SHAM+ND group. Two-way ANOVA indicated significant association of FFA concentrations with diet, while insulin concentrations showed significant association with both ovariectomy and diet. A significant interaction was observed between ovariectomy and diet.
Liver SREBP1c and FAS mRNA expression
Liver SREBP1c and FAS are involved in regulation of lipogenesis. The relative expression of SREBP1c was increased in the OVX+HD group, although without statistical significance (Table 6). In addition, significantly higher FAS expression was observed in the OVX+HD group. These results indicate that the coexistence of OVX and HD affected the expression of lipogenic genes. However, no statistically significant interaction was found between OVX and diet.
Tumor tissue IR, AKT, mTOR, and AMPK expression
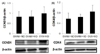
To determine the molecular mechanisms responsible for tumor growth, expression of IR-mediated signaling molecules including AKT and pAKT was measured in mammary tumor tissues. Significantly higher tissue protein expression levels of IR were observed in the OVX+HD group compared to the SHAM+ND and SHAM+HD groups (Fig. 1A), and significantly higher pAKT/AKT levels were observed in the OVX+HD group compared to the SHAM+ND group (Fig. 1B). Significantly higher protein expression of mTOR was observed in the OVX+HD group, while significantly lower expression of pAMPK/AMPK was observed in the OVX+HD group compared to expression in other groups (Fig. 1C and 1D).
Cell cycle regulation
Activation of AMPK and mTOR was involved in regulation of the cell cycle, including cyclin D1 (CCND1) and CDK4 protein. Elevated expression levels of CCND1 and CDK4 were observed in the OVX+HD group compared to other groups, but without statistical significance (Fig. 2A, B).
DISCUSSION
Despite an inverse relationship between BMI and breast cancer risk in premenopausal women, obesity is a recognized risk factor in postmenopausal breast cancer development [27]. Although it is presumed that the interaction between obesity and breast cancer risk is altered by the availability of estrogen, evidence supporting the association between dietary components including dietary fat and the risk of breast cancer is limited. Because estrogen is known to regulate fat metabolism, including lipogenesis and lipolysis, it is presumable that interactions between dietary fat intake, energy metabolism, and estrogen availability play a critical role in mammary tumor development.
Estrogen deprivation often leads to diminished insulin sensitivity, possibly due to accelerated fat accumulation. Previous studies have shown that estradiol repletion can overcome high-fat diet-induced or FFA-induced insulin resistance [2829]. Accordingly, high circulating concentrations of insulin in postmenopausal women may be a possible connection between postmenopausal obesity and breast cancer risk [30].
However, a limited number of studies have examined the interactive effects of estrogen deprivation and excess dietary energy supply on mammary tumor development. Genetically obese ovariectomized Zucker rats showed higher susceptibility to DMBA-induced mammary tumor development compared to sham-operated rats [31]. However, in another study high-fat diet stimulated mammary tumor development was not ovarian-dependent [32]. In this study, we evaluated the effects of estrogen deprivation on breast tumor growth in mice fed either a 45% fat diet or 15% fat diet. The animal model of DMBA-induced mammary gland tumors was used because this carcinogen has been implicated in mammary tumorigenesis that is histologically similar to hormone-dependent human breast adenocarcinomas [33]. Results showed that there was no difference in body weight among experimental groups. Previous studies have shown that body weight of C3H/HeN mice fed HD was higher than that of ND supplemented C3H/HeN mice [3435]. In many other studies, OVX rodents became obese rapidly with body fat accumulation [123637]. However, mice treated with DMBA had significantly lower final body weights, body fat weights, and carcass energy contents compared to mice that received the corn oil placebo [38]. Another study also reported that DMBA treatment resulted in suppression of body weight gain of the animals [39] possibly due to rapid growth of tumor tissues, which accelerates tissue wasting, indicating that the absence of body weight gain in animals with either high-fat feeding or OVX might be due to tumor-induced tissue wasting. Estrogen depletion is known to be associated with dysregulation of lipid metabolism, which may contribute to the accumulation of intra-abdominal fat in postmenopausal women [40]. Estradiol reduces fatty acid and triglyceride synthesis through the down-regulation of lipoprotein lipase and fatty acid synthase [12]. We have determined the liver tissue expression of SREBP1c and FAS, both of which are involved in lipogenesis. Results showed higher liver tissue expression of SREBP1c and FAS in the OVX+HD group compared to the other groups, although no statistical significance was found in SREBP1c expression. The fact that cancer cachexia and tissue wasting are often associated with decreased insulin sensitivity and hyperglycemia has also been pointed out [41], indicating that tumor growth and hypercatabolism can lead to hyperinsulinemia.
Estradiol depletion is associated with insulin resistance in humans and rodents [293042]. The increased circulating concentration of FFA in OVX rodents contributes to development of insulin resistance [4344]. Postmenopausal insulin resistance is also associated with an elevated blood concentration of inflammatory markers, TG, LDL-cholesterol, and FFA [4546]. Our study results clearly showed significantly higher concentrations of circulating insulin in the OVX+HD group than other groups.
In addition, the levels of IR increased significantly in tumor tissues of the OVX+HD group. Because insulin has been suggested as a key molecule to explain obesity-related cancers including colon and breast, we hypothesized that the larger tumor size in OVX+HD may be mediated through the higher availability of insulin as well as the up-regulation of IR in tumor tissues. Another recent study reported that tumor weight was significantly higher in OVX mice fed a high-fat diet, compared to OVX mice fed a low fat diet in a mouse breast cancer model [47]. In addition, obese mice inoculated with mouse mammary tumor virus (MMTV) Wnt-1 mammary tumor cells did not exhibit tumor growth in the presence of estrogen; however, in the absence of the ovaries, obese mice exhibited higher Wnt-1 tumor growth [48]. These results suggest a strong association of excess energy intake with tumor growth in the absence of ovarian estrogen. Therefore, it is presumable that excess energy intake of postmenopausal women may pose greater risk of developing tumors compared to that of premenopausal women.
The tumor incidence was 75% in the SHAM+HD group, 57.1% in the SHAM+ND group, 33.3% in the OVX+ND group, and 42.9% of the OVX+HD group, indicating no clear association between the number of tumors and OVX or dietary energy. Although further investigations are necessary, the higher concentrations of circulating insulin due to OVX and the excess energy supply may not be directly associated with DMBA-induced cancer initiation. Because only two animals developed tumors in the OVX+ND group, no statistical comparisons for tumor tissue biomarkers of cell growth were made with animals in this group. It was reported that breast tumor incidence in Zucker rats, where 30% of the lean sham-operated group, 59% of the obese sham-operated group, 0% of the lean ovariectomized group, and 36% of the obese ovariectomized group developed mammary tumors [31], implying that DMBA-induced mammary tumor formation is dependent on estrogen levels as other studies have also indicated [495051]. However, it has been suggested that estrogen synthesized from adipose tissue is sufficient to promote mammary tumor development [52]. In this study, since OVX+ND mice were shown to have little adipose tissue, the level of systemic and local estrogen production may not be sufficient for development of tumors. In another study examining incidence of mammary cancer induced by DMBA combined with different concentrations of estradiol in 8-week-old Sprague-Dawley rats, it was found that mammary tumors appear for the first time between the 12th-17th week [28]. Fifty percent of mice had tumors by the 36th, 19th, and 18th week, with estradiol doses of 1, 2, and 3 mg, respectively. At the 36th week, the incidence rate of breast tumors was 50%, 73%, and 100%, for estradiol doses of 1, 2, and 3 mg, respectively. These results imply that estrogen plays an important role in the initiation of mammary tumor formation induced by DMBA. In our study, OVX was performed at 4 weeks, which may have caused depletion of circulating estrogen required for initiation of carcinogenesis.
Hyperinsulinemia can indirectly affect tumorigenesis through activation of insulin/AKT signaling. A previous study reported that larger tumor size in obese women is the result of growth stimulation, and IR-mediated AKT signaling is a major growth pathway [53]. Another study indicated that IR was overexpressed in human breast cancer cells [54]. To investigate involvement of IR-mediated AKT pathway signaling in mammary tumor growth, we measured protein expression levels of IR and AKT in tumor tissues. IR protein level was significantly higher in the OVX+HD group compared to the other groups, and AKT phosphorylation and mTOR expression were also significantly higher in the OVX+HD group. mTOR, which regulates essential cell growth signals through cell cycle progression, is an important downstream effector of AKT [55]. A previous study reported that in breast cancer, activation of the AKT/mTOR pathway is responsible for cell survival [56]. Therefore, it is presumable that IR-mediated AKT activation is responsible for the larger tumor size in the OVX+HD group. Epidemiological evidences have suggested that excess body fat is a risk factor for development of breast cancer in postmenopausal women, but not in premenopausal women [23]. Results from this study showing an interactive effect of high-fat diet and OVX on circulating concentration of insulin support higher breast cancer risk among postmenopausal women with excess body weight.
We found that OVX+HD suppressed the activation of AMPK, which is potentially associated with tumor cell growth. Interestingly, it is reported that the levels of AMPK phosphorylation are reduced by OVX and excess fat mass [5758]. In addition, AMPK is known as a major tumor suppressor kinase that acts through p53-dependent cell cycle regulation [5960]. AMPK is also known to suppress cell proliferation through down regulation of mTOR [61]. Metformin, a well-known AMPK activator, has been consistently shown to inhibit breast cancer cell growth [56]. In this study, we found that the OVX+HD group showed higher AMPK expression, which may have led to down-regulated expression of mTOR, thereby suppressing tumor growth. Estrogen has been implicated in maintenance of insulin sensitivity [62], and stimulated AMPK phosphorylation by17β-estradiol through estrogen receptor α in 3T3-L1 adipocytes has also been reported [63]. In addition, injections of estradiol resulted in activation of AMPK in ovariectomized mice [64]. Therefore, it is possible that the significantly larger tumor size observed in the OVX+HD group resulted from the combined effects of OVX and excess energy supply. A previous study reported that mTOR facilitates CCND1, which is required for the G1 to S phase transition of the cell cycle [65]. We measured protein expression of CCND1 and CDK4 as downstream targets of mTOR in mammary tumor tissue. Although expression levels of CCND1 and CDK4 were not significantly different, the OVX+HD group tended to show higher expression levels of cell cycle-controlling molecules.
In conclusion we demonstrate that a high-fat diet in OVX animals leads to development of insulin resistance, which may accelerate mammary tumor growth through the IR-mediated AKT pathway and inactivation of AMPK in vivo. The current study clearly showed that dietary fat induces systemic insulin resistance and mammary tumor growth in estrogen-deprived animals in the absence of body weight gain. High circulating insulin in combination with increased IR in tumor tissues may result in stimulation of AKT/mTOR signaling and inactivation of AMPK leading to the acceleration of solid tumor growth. The current results suggest that a high-fat diet can stimulate breast cancer progression in postmenopausal women even those maintaining normal BMI.







 XML Download
XML Download