

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Soybean, an important plant protein source for human and animals, has been reported to have beneficial activities, including antiobesity, antdiabetic, anti-inflammatory, and anti-cardiovascular disease properties [1]. Fermentation is an excellent processing method for improving nutritional and biological properties of soybean. Previous studies have shown that fermented soybean foods contain higher level of bioactive compounds [23] and exhibit stronger anti-oxidative [4567] and anticancer activities [89] than unfermented ones. These results have drawn attention to the potential preventive role of fermented soybean foods against many chronic diseases.
Doenjang, a Korean long-term fermented soybean paste, is usually consumed as an ingredient or seasoning in various sauces, soups, or stews. It is regarded as a good source of essential amino acids and fatty acids, particularly for people on a grain and vegetable based diet. Doenjang has been manufactured every year for centuries at homes in Korea using traditional methods, which employ Bacilli subtilis, Bacillus licheniformis, and molds such as Rhizopus, Mucor and Aspergillus oryzae from rice straw and local environments [1011]. However, with rapid urbanization and industrialization of society, manufacture by inoculation of some major microorganisms is increasing with shorter fermentation time and aging under controlled conditions in a factory.
It was reported that that Doenjang intake resulted in significant reduction of body weight gain [121314], visceral adipocyte content and circumference [12], serum oxidative stress and proinflammatory cytokine levels in obese rats fed a high fat diet [4,13]. Inflammation is a primary host defense reaction, and considered a beneficial and necessary attempt of the organism to eliminate the aggressive agent and to start the healing process against many pathological conditions. There is convincing evidence of a close association between proinflammatory stress and metabolic stress in obesity and aging-related chronic diseases such as atherosclerosis and insulin resistance [15]. When the inflammation control system does not function properly and chronic inflammation persists, development of diseases including cancer may occur [1617]. Therefore, pharmacological control of the many aspects of inflammation continues to be a major issue in the development of new drugs [18].
Overproduction of reactive oxygen species (ROS) can cause oxidative damage to membrane lipids, DNA, proteins, and lipoproteins, eventually leading to development of chronic diseases including cancer, chronic inflammation, and cardiovascular diseases. ROS is a stimulator of the inflammatory response via activation of signaling pathways including nuclear factor kappa B (NF-κB) [19].
Most isoflavones in raw, cooked, or unfermented soybean exist in the glycoside form. However, cleavage of β-glycosyl bond of isoflavone glycoside by β-glucosidase released from rapidly growing microorganisms during the process of fermentation results in higher contents of isoflavone aglycones, including genistein and daidzein [20212223]. Genistein, a major isoflavone aglycone in fermented soybean products, is reported to have various biological functions such as preventive coronary heart disease and osteoporosis, as well as anti-oxidant, anti-neoplastic, and anti-inflammatory compounds [1524]. In terms of anti-inflammatory activity, genistein significantly reduced the production of nitric oxide (NO) and prostaglandin (PG) E2 by suppression of inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2 expression at the transcriptional level in lipopolysaccharide (LPS)-treated RAW 264.7 macrophages [25]. Genistein also effectively inhibited the LPS-induced overproduction of proinflammatory cytokines such as tumor necrosis factor (TNF)-α and interleukin (IL)-6 in a dose dependent manner via inhibition of NF-κB activation following adenosine monophosphate-activated protein kinase (AMPK) stimulation [26].
However, few studies have reported on antioxidant and anti-inflammatory activities of Doenjang during different stages of aging. In this study, we investigated the changes in antioxidant and anti-inflammatory activities of Doenjang during aging.
MATERIALS AND METHODS
Sample preparation, extraction, and fraction
Doenjang was manufactured by SunchangJangryu Co., located in Sun Chang County in Jeonlabuk-do, Korea. The preparation method is shown in Fig. 1.
For our experiment, parts of steamed soybean (SB), Meju (MJ), and Doenjang aged for 0, 1, 3, 6, 8, and 12 months (D0, D1, D3, D6, D8, and D12) were sampled. Samples were freeze-dried (Samwon, Sungnam, Korea), powdered, and stored at -20℃. A 100 g portion of each dried sample was extracted with ethanol (v/v) 10 times with stirring for 24 h. The supernatant was removed and filtered with Whatman paper (No. 2), and the residue was extracted once more with ethanol and filtered. Two filtrates were combined, and evaporated for removal of ethanol using a rotary vacuum evaporator (EYELA, Tokyo, Japan), then concentrated ethanol extract was dissolved in distilled water and sequentially fractionated with hexane (Hx), dichloromethane (DM), ethylacetate (EA), and n-butanol (BT), leaving an aqueous layer (DW). All fractions were also concentrated, freeze-dried, dissolved in DMSO or distilled water at 100 mg/mL, and stored at -20℃.
Determination of total phenolic and flavonoid contents
For determination of total phenolic and flavonoid contents, 1.0 g of each dried sample powder was stirred in 50 mL of 80% ethanol for 24 h at room temperature and filtered with Whatman paper (No.2). Total phenolic concentration in the filtrate was determined according to Singleton et al. [27]. Briefly, 100 µL of samples was mixed with 1 mL of 0.2 N Folin-Ciocalteau reagent and 200 µL of 20% sodium carbonate, and stood in the dark for 1 h. The absorbance was read at 760 nm by spectrophotometer using tannic acid as a standard. Total flavonoid content was determined according to the modified AOAC method [28]. Briefly, 100 µL of samples was mixed with 900 µL of 90% diethylene glycol and then 20 µL of 1 N NaOH, and incubated in a 37℃ water bath for 1 h. The absorbance was read at 420 nm by spectrophotometer using rutin as a standard.
Isoflavone analysis
Each ethanol extract was dissolved in 70% ethanol, centrifuged at 10,000 × g for 5 min, and the supernatant was filtered with a0.45 µm membrane (PVDF syringe filter, Whatman, Germany). Reversed-phase HPLC system (JASCO, Japan) with an ODS A303 column (25 µm, 4.6 × 250 mm, YMC, USA) was used for isoflavone analysis. A linear HPLC gradient was composed of (A) 0.1% acetic acid in acetonitrile and (B) 0.1% acetic acid in water. The solvent flow rate was 1.0 mL/min. Following injection of 20 µL of sample, eluted isoflavones were detected by the absorbance at 254 nm on a UV detector. Quantitative data for daidzin, glycitin, and genistin and their malonyl, acetyl, and aglycone forms were obtained by comparison with 12 known standards. Isoflavone standards were obtained as follows: genistein, daidzein, and glycitein from Sigma (St. Louis, MO, USA), genistin, daidzin, and glycitin from Indofine (Hillsborough, NJ, USA), and malonylgenistin, malonyldaidzin, malonylglycitin, acetylgenistin, acetyldaidzin, and acetylglycitin from LC Labs (Woburn, MA, USA). All data are presented as mean of two measurements.
Measurement of 2,2-diphenyl-1 picryl hydrazyl (DPPH) radical scavenging activity
DPPH radical scavenging activity was determined as previously described [29]. The various concentrations of each sample in 10 µL were mixed with 190 µL of 200 µM DPPH in ethanol, incubated at 37℃ for 30 min, and their absorbance was read at 517 nm by spectrophotometer. The reduction in absorbance is proportional to the ability of the sample to eliminate DPPH radicals. Ascorbic acid was used as a positive control. DPPH radical scavenging effect was calculated as follows; DPPH radical scavenging activity (%) = [(Acontrol - Asample)/Acontrol] × 100. IC50 value, the sample concentration needed to reduce 50% of DHHH radicals, was calculated from the equation between sample concentration and DPPH scavenging activity.
Measurement of Ferric reducing antioxidant power (FRAP)
The method described by Yildrim et al. [30] was used for determination of FRAP. Various concentrations of sample in 0.2 M phosphate buffer (pH 6.6), 1% potassium ferricyanide were incubated in a water bath at 50℃ for 30 min, followed by addition of 10% trichloroacetaic acid. Following centrifugation of solution at 3000 rpm for 10 min, supernatant was collected, mixed with 0.1% FeCl3 and the absorbance read at 700 nm by spectrophotometer. The result was expressed as µg/mL of ascorbic acid, positive control.
Cell culture and sample preparation
RAW 264.7 murine macrophage cells were cultured in DMEM and supplemented with 10% FBS, 2 mM glutamine, 100 unit/mL penicillin, and 50 µg/mL streptomycin at 37℃ in a cell incubator of 5% CO2. Fractionated samples were dissolved in DMSO (100 mg/mL), filtered by sterile 0.2 µm syringe filter, and stored at -20℃.
Cell viability
Cell viability was determined by MTT assay [31]. RAW 264.7 cells were seeded in 96-well plate at a density of 5 × 103 cells/well. Following 24 h incubation in 10% FBS containing DMEM in a cell incubator, culture medium was replaced with serum-free medium including various concentrations of sample. Following further 24 h incubation, medium was replaced with 180 µL of serum-free and 20 µL of 5 mg/mL MTT solution. Following 4 h incubation at 37℃, the supernatant was discarded and 200 µL of DMSO was added to dissolve the formazan crystals formed in the cells. The absorbance was read at 540 nm using a microplate reader (Bio-Tech Instrument, Winooski, VM, USA). The results were expressed as percentage of control.
Measurement of NO, PGE2, IL-6, and TNF-α production
RAW 264.7 macrophage cells were seeded in 12-well plate at a density of 5 × 105 cells/well in 2 mL of DMEM containing 10% FBS, and incubated in a cell incubator for 24 h. Medium was replaced with 1% FBS and phenol red-free medium including various concentrations of sample, incubated for 2 h, and then LPS (1 µg/mL) was applied except negative control. Following further 22 h incubation, NO, PGE2, IL-6, and TNF-α concentrations were measured in cultured medium. NO concentration was measured by Griess reaction [32]. Each 100 µL of cultured medium was mixed with 100 µL of Griess reagent and incubated at room temperature. After 20 min, the absorbance was read at 540 nm using an ELISA plate reader. NO concentration was calculated from a standard curve drawn with known concentrations of sodium nitrite. PGE2, IL-6, and TNF-α concentrations were determined using ELISA kits (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
Western blot analysis
Cells were washed with PBS and lysed with RIPA buffer (Sigma, Saint Louis, MS, USA) and protease and phosphatase inhibitor cocktail (Roche, Germany) on ice for 20 min. The cell debris was pelleted by centrifugation at 10,000 g for 15 min. The protein level of the supernatant was determined using Bradford method [33]. Total cell lysates were resolved in sodium dodecyl sulfate (SDS) sample buffer (Sigma, Saint Louis, MS, USA), separated by electrophoresis on 10% SDS polyacrylamide gel in the Mini-PROTEIN system (BIO-Rad) and transferred onto Immobilon membranes (Millipore, Bedford, MA). The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline containing 0.1% Tween-20 (TBST, pH 8.0) at room temperature for 1 h, and probed with rabbit polyclonal antibodies against iNOS, COX-2, or anti-β-actin monoclonal antibody (Cell signaling, Technology Inc, OR, USA) diluted in TBST with 5% nonfat milk with overnight shaking at 4℃. After washing with TBST, the membranes were treated with secondary antibody (horseradish peroxidase-labeled IgG, 1:1000-1:2000) for 1 h at room temperature and washed. Protein bands were visualized by ECL solution (Pierce, Rockford, IL) and quantified using Luminescent Image Analyzer (FujiFilm, Japan).
RESULTS
Contents of total phenolics and flavonoids in dried Doenjang
Contents of total phenolics and flavonoids in freeze-dried SB, MJ, and Doenjang are shown in Table 1. Total phenolic content was significantly increased by fermentation, and further increased by aging (P < 0.05). Different from total polyphenol content, total flavonoid contents in SB were not significantly changed by dry fermentation, slightly dropped in D0 and D1, and then recovered after 6 months of aging.
Antioxidant activity of ethanol extract
DPPH radical is a stable free-radical donor widely used for estimation of free-radical scavenging activity. It is reduced in the presence of an antioxidant molecule, producing a color change from violet to yellow. DPPH radical scavenging activities (IC50) of the samples are shown in Table 1. DPPH radical scavenging activity was rapidly increased (P < 0.05) by the fermentation and 1 month of aging, however, it was not significantly increased after 1 month of aging. IC50 was decreased from 21.0 mg/mL in SB to 15.5 mg/mL in MJ, 4.5 mg/mL in D1 and 3.9 mg/mL in D12. Total phenolic content (r = -0.8772, P < 0.001), aglycone isoflavone content (r = -0.9195, P < 0.001), and glycoside isoflavone content (r = 0.9595, P < 0.001) showed significant correlation with IC50 value for DPPH radicals, while no significant correlation was observed between total flavonoid content and DPPH radical scavenging activity (data not shown).
FRAP was determined at various concentrations of ethanol extract, 500, 1000, and 2000 µg/mL (Table 2). At all concentrations, FRAP in SB and MJ did not differ from each other, indicating that FRAP was not increased by dry fermentation. However, FRAP was rapidly increased by wet fermentation and aging (P < 0.05). At 2000 µg/mL, FRAP in MJ, 10.2 µg AA eq./mL, increased up to 19.9 µg AA eq./mL in D0, 32.1 µg AA eq./mL in D1 and 53.5 µg AA eq./mL in D12.
Total phenolic and flavonoid contents and antioxidant activity in different fractions
Total phenolic contents in DM and EA fractions were 330.35 mg TA/mg and 269.24 mg TA/mg, respectively, which were significantly higher than those in Hx, BT, and DW fractions (P < 0.05). Total flavonoid contents in DM and EA were 32.69 mg RT/mg and 32.02 mg RT/mg, respectively, which were also significantly higher than those in the other fractions (P < 0.05) (Table 3).
DPPH radical scavenging activities of DM and EA fractions were significantly higher (P < 0.05) than those of the other fractions. At 100 and 200 µg/mL, DPPH radical scavenging rate was 59.3% and 88.1% in DM fraction, and 80.9% and 94.9% in EA fraction, respectively (Table 3).
Isoflavone content
Three major isoflavone contents in samples are shown in Table 4. Total isoflavone content of diadzein, glycitein, and genistein showed a gradual decrease during fermentation and the aging process. Overall, in examination of the isoflavone composition, aglycone content and its proportion were increased by fermentation and aging. In detail, however, aglycone content peaked in D3, and its proportion peaked in D6, and then both of them showed a slight decrease until 12 months of aging, while glycoside content and its proportion were gradually decreased until 6 months of aging and then showed a slight increase until 12 months of aging.
Effect of DM and EA fractions of Doenjang on NO and PGE2 production in RAW 264.7 macrophages
DM and EA fractions of SB, D0, D6, and D12 did not show cytotoxicity to RAW 264.7 cells at the treated concentrations (10, 25, 50, and 100 µg/mL) as shown in Fig. 2. To investigate potential anti-inflammatory activity of Doenjang, the RAW 264.7 macrophages, which produce NO and PGE2 upon stimulation with LPS, were treated with DM and EA fractions of steamed soybeans and Doenjang samples (D0, D6, and D12), and the accumulated NO and PGE2 concentrations in cultured medium were measured. LPS (1 µg/mL) treatment resulted in significantly increased production of NO and PGE2, as shown in Fig. 3.
DM fractions of SB and D0 did not suppress the NO concentration at all treated concentrations. However, DM fraction of D6 significantly suppressed (P < 0.05) the NO concentration to 88.8% of LPS-treated control at 100 µg/mL, and DM fraction of D12 significantly suppressed (P < 0.05) the NO concentration to 83.3% and 55.6% of LPS-treated control at 50 and 100 µg/mL, respectively. DM fraction of SB did not influence the PGE2 concentration, while DM fraction of D0, D6, and D12 significantly suppressed (P < 0.05) it at 50 and 100 µg/mL in a dose dependent manner. At the highest concentration (100 µg/mL), DM fractions of D0, D6, and D12 suppressed the PGE2 concentration to 71.0%, 62.9%, and 25.0% of LPS-treated control, respectively.
EA fractions of SB and D0 did not suppress the NO concentration. However, EA fraction of D6 significantly suppressed (P < 0.05) the NO concentration to 89.3% and 84.6% of LPS-treated control at 50 and 100 µg/mL, respectively, and EA fraction of D12 significantly suppressed (P < 0.001) it to 74.3% and 52.5% of LPS-treated control at 50 and 100 µg/mL, respectively. EA fraction of SB did not influence the PGE2 concentration, while EA fractions of D0 and D6 significantly suppressed (P < 0.05) it at 100 µg/mL and EA fraction of D12 significantly suppressed (P < 0.001) it at 50 and 100 µg/mL. At 100 µg/mL, EA fractions of D0, D6, or D12 suppressed the PGE2 concentration to 68.7%, 74.4%, and 28.3% of LPS-treated control, respectively. DM and EA fractions of D12 commonly showed significantly higher (P < 0.05) suppression effects on NO and PGE2 production at 100 µg/mL than those of SB, D0, and D6.
Effect of DM and EA fractions of Doenjang on IL-6 and TNF-α production in RAW 264.7 macrophages
To investigate potential anti-inflammatory activity of Doenjang, the accumulated IL-6 and TNF-α concentration in cultured medium was also measured. LPS (1 µg/mL) treatment resulted in significantly increased production of IL-6 and TNF-α, as shown in Fig. 4.
DM fractions of SB, D0, and D6 did not suppress the IL-6 and TNF-α concentration at all treated concentrations, while DM fraction of D12 significantly suppressed (P < 0.05) the IL-6 and TNF-α concentrations to 70.6% and 86.9% of the LPS-treated control at 100 µg/mL.
Comparing the effects of EA fractions on IL-6 production at 100 µg/mL, D0, D6, and D12 significantly suppressed (P < 0.05) the IL-6 concentration to 80.9%, 89.3%, and 40.1% of the LPS-treated control, respectively, while SB did not. In addition, EA fraction of D12 significantly suppressed (P < 0.05) the IL-6 concentration in a dose-dependent manner to 81.5%, 77.3%, and 40.1% of the LPS-treated control at 10, 50, and 100 µg/mL, respectively. EA fractions of all samples including SB significantly suppressed (P < 0.05) the TNF-α concentration to 91.2%, 90.4%, 89.2%, and 86.8% of the LPS-treated control at a low concentration of 10 µg/mL, respectively, however, significant suppression was not observed at 50 and 100 µg/mL.
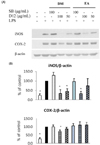
Effect of DM and EA fractions of Doenjang on iNOS and COX-2 protein expression in RAW 264.7 macrophages
Because DM and EA fractions of D12 suppressed NO and PGE2 production in LPS-treated RAW 264.7 cells, Western blot analysis was performed to examine whether those fractions of D12 inhibited iNOS and COX-2 protein expression.
As shown in Fig. 5, extremely low detectable levels of iNOS and COX-2 proteins were expressed by the cells under the basal condition, however, iNOS and COX-2 protein expression was remarkably increased by LPS treatment. Treatment with DM fractions of D12 resulted in reductions of iNOS protein expression to 44.2% and 33.9% of the LPS-treated control at 50 and 100 µg/mL, respectively, and COX-2 protein expression was reduced to 87.1% and 71.6%, respectively, while DM fraction of SB did not reduce iNOS and COX-2 expression. Treatment with EA fraction of D12 resulted in reduction of iNOS protein expression to 75.3% and 32.7% of the LPS-treated control at 50 and 100 µg/mL, but COX-2 expression was not reduced.
DISCUSSION
ROS are responsible for damage to phospholipids in biomembranes by lipid peroxidation, and are known to be one of the stimulators of the inflammatory response [193435]. Strong association of the ability to scavenge the DPPH radical with lipid peroxidation has been reported [36] and relatively high positive correlation between anti-inflammatory and antioxidant activity of plant extracts was observed [373839].
Several studies found that phenolic compounds were responsible for the antioxidative activity in fruits, vegetables, and grains [404142]. Soybeans contain natural antioxidants. Tocopherol and phosphatides are found in soybean oil, while the non-oil component contains many isoflavones, a class of phytochemicals. Isoflavonoids from legumes exist mainly in glycoside conjugates that are less bioavailable [4344]. The combined isoflavone aglycone and phenolic acids were reported to account for nearly all in vitro antioxidant activity of soybean and soy products [74546]. Dsiedzic et al. [47] reported that phospholipids rich in soy product can synergistically promote antioxidant activity of isoflavone.
Chai et al. [3], who reported that IC50 for DPPH radical scavenging activity was soybean > Meju > Doenjang > Ganjang and was negatively correlated with total polyphenol content, total flavonoid content, and total protein content, suggested that vanillic acid and protocatechuric acid are the effective compounds of higher antioxidant activity of soybean fermented foods. Melanoidin produced during fermentation of Doenjang also contributed to increased antioxidant activity. Browning of Doenjang is the result of enzymatic and nonenzymatic processes. Enzymatic browning is known to be mainly due to melanin produced by tyrosinase released from Bacillus, and nonenzymatic browning is mainly due to Maillard reaction of amino acid, sugars and carbonyl substance in Doenjang [48]. Kim et al. [49] already reported that melanoidin content was high in chloroform extract of Meju and methanol extract of Doenjnag. Jeong et al. [50] isolated Lactobacillus from traditional Meju showing high DPPH radical scavenging activity. In addition, 7.3',4-trihydroxyisoflavone, 7,8,4'-trihydroxyisoflavone, and 6,7,4'-trihydroxyisoflavone, isolated in long-term fermented and aged traditional Doenjang, was reported to show strong activity for scavenging DPPH radical and superoxide [5152].
In this study, total isoflavone content showed a gradual decrease with increase of processing time from 3.01 mg/g in SB to 1.46 mg/g of MJ, 1.23 mg/g in D0 and 0.74 mg/g in D12. The reason could not be clearly explained, however, it could be due in part to hydrolysis and release out from the solid part of Meju into the liquid part during wet fermentation in saline and chemical biotransformation during the aging process. Wang and Murphy [53] reported that 61% and 44% of isoflavones in soybeans were lost in temphe and tofu, respectively.
Many researchers reported that aglycone isoflavone content and its proportion of the total isoflavone content was increased by fermentation [212223]. In this study, total isoflavone content showed a gradual decrease with increase of time during fermentation and aging, however, aglycone content and its proportion were increased until 3 or 6 months of aging, and then slowly decreased until 12 months of aging. High β-glucosidase activity of Aspergillus oryzae and Bacillus licheniformis, used in this study, was reported [5455]. In this study, it was observed that not only β-glucoside isoflavone but also malonyglucoside and acetylglucoside contents were decreased by fermentation, thereby, the accumulated aglycone content increased by fermentation and aging, however, it reached the maximum level at 3 months of aging and then decreased slowly.
It was questionable why aglycone content in Doenjang decreased after 3 months of aging, and the antioxidant activity was sustained or rather increased up to 12 months of aging despite a decrease in aglycone content after 3 months of aging. Probable reason might be due to biotransformation of aglycones into hydroxyisoflavones, such as 8-hydroxydaidzein, hydroxygenistein, and hydroxyglycitein [53]. Lee et al. [55] recently reported that fermentation of a solution of ground steamed soybeans with Aspergillus oryzae at 30℃ decreased acetylglycoside and glycoside content and aglycone content increased after the first 12 hours of fermentation, however, aglycone content was decreased after the second or third 12 hours of fermentation due to the conversion to hydroxyisoflavone, and the content of this substance showed strong correlation with antioxidant activity.
It is needed to age the initial Doenjang for food. It is usually sold after 3 months of aging following the fermentation process, and consumed for a long time approximately several months to years. The limitations of our study are that it was not sampled at over 12 months of aging with only one sample at each time. In addition, the aglycone isoflavone content was not measured in the different fractions of Doenjang ethanol extract.
In this study, it was observed that DM and EA fractions from Doenjang ethanol extract showed higher antioxidant activity and polyphenol content than the other fractions, and especially these two fractions of D12 extract showed the highest activity among samples. Therefore, we performed further experiments on anti-inflammatory activity of Doenjang using DM and EA fractions of D12 ethanol extract in LPS-activated RAW 264.7 macrophages.
Macrophage plays an important role in the inflammatory response. When activated, macrophages release NO, cytokines, and lipid mediators such as prostaglandins, which promote inflammation by stimulating cellular migration to the target site [15]. LPS, the major component of the outer membrane of Gram-negative bacteria, consists of a lipid and a polysaccharide joined by a covalent bond. LPS activates a number of major cellular pathways that play critical roles in the pathogenesis of inflammatory response [56].
NO is an important physiological messenger in many biological systems including immunological, neuronal, and cardiovascular tissues [5758]. PGE2, a key inflammatory mediator, is converted from free arachidonic acid by cyclooxygenase [57]. During the inflammatory process, significant quantities of the pro-inflammatory mediators of NO and PGE2 were generated by iNOS and COX-2, respectively. COX enzyme possesses both cyclooxygenase and peroxidase activities. Prostaglandins produced by COX impair immune surveillance and modulate proliferation in a variety of cell types. The peroxidase function contributes to activation of carcinogens. High production of NO has been shown to cause DNA damage as well as mutations during infection and inflammation. Over-expression of either COX-2 or iNOS may be intimately involved in the pathogenesis of many diseases, such as colon cancer, multiple sclerosis, neurodegenerative diseases, and heart infarction [25].
NF-κB, activator protein-1, CCAAT/enhancer-binding protein and cyclic-AMP-response element-binding protein are commonly or individually involved in regulation of inflammatory genes including iNOS and TNF-α [59]. In particular, activation of NF-κB induces expression of various inflammatory target genes involved in inflammation, inducing iNOS, COX-2, IL-6, and TNF-α [60]. One study [56] reported that genistein blocked LPS-induced intracellular inflammatory signaling cascades by suppressing ROS production and NF-κB signaling pathway. In addition, genistein potently suppressed binding of LPS to toll-like receptor 4 on BV2 microglial cell surface.
In this study, DM fraction of D12 extract obviously suppressed COX-2 protein expression while EA fraction of D12 extract did not, even though these DM and EA fractions markedly suppressed PGE2 production to a similar extent in LPS-activated macrophages. It is considered that suppressed PGE2 production by EA fraction may be due to inhibition of COX-2 activity rather than expression of COX-2 at the protein level, and direct suppression of microsomal PGE2 synthase (mPGE2) activity or inhibition of 15-OH PG dehydrogenase activity, converting PGs into inactive forms. Koeberle et al. [61] reported that curcumin suppressed PGE2 production via inhibition of COX-2 expression and direct activation of mPGE2 activity.
Increased total polyphenol content and/or isoflavone aglycone during fermentation is known to be associated with various bioactivities [715242745]. Results of this study indicated that effective substances of antioxidant and anti-inflammatory activities of Doenajng could be phenolics, aglycone isoflavone and some other substances dissolved rich in DM and EA fractions. Taken together, it is concluded that 12 month-aged Doenjang has higher anti-inflammatory and antioxidant activities than soybeans or Doenjang aged for a shorter time, and can be used as a beneficial food for human health.
Further study is needed to investigate the underlying biomolecular mechanisms for anti-inflammatory activity of Doenjang in various systems, and to find an effective candidate substance dissolved in DM and/or EA fractions.








 XML Download
XML Download