

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Alcoholic liver disease (ALD) is a major cause of mortality and morbidity worldwide [1]. The liver is the main organ affected because it is the major site of alcohol metabolism and produces toxic metabolites from alcohol, such as, acetaldehyde, acetate, and reactive oxygen species [2]. The oxidative metabolism of alcohol produced acetaldehyde, which contributes to cell and tissue damage and increased oxidative stress [3], and increased oxidative stress decreases antioxidant enzymes and glutathione (GSH), resulting in DNA damage and the necrosis and apoptosis of hepatic cells [4].
Alcohol-induced oxidative stress promotes inflammation, which is aggravated by an increase in pro-inflammatory cytokines levels and by the up-regulation of the inflammatory cascade [5]. Circulating levels of the endotoxin, lipopolysaccharide (LPS), increase in rodents on a chronic alcohol diet [6]. Furthermore, LPS recognition by toll-like receptor 4 (TLR4) results in the recruitments of adaptor molecules that activate nuclear factor kappa B (NF-κB), which increases the productions of pro-inflammatory cytokines, such as, 'tumor necrosis factor α (TNF-α)' and 'interleukin 6 (IL-6)' [6]. In particular, these two cytokines are the principle mediators of early alcohol-induced liver injury. Treatment of ALD is accomplished using anti-inflammatory agents, antioxidants, and agents directed against the progression to fibrosis [7]. However, owing to the adverse side effects associated with many agents, alternative natural therapeutics are needed.
Umbelliferone (UF; 7-hydroxycoumarin; Fig. 1) is present in fruits and roots plants, such as, the golden apple, the bitter orange, and carrot [8,9,10], and several studies have shown that UF can exert potent antioxidant, antidiabetic and antitumor effects [11,12]. Kassim et al. [13] demonstrated the antioxidant property of UF by the 1,1-diphenyl-2-picrylhydrazyl free radical scavenging, and Ramesh and Pugalendi [11,14] have reported that UF has significant glucose reducing and antioxidant properties, as demonstrated by reductions in gluconeogenic enzyme levels and lipid peroxidation. In a previous study, we found that UF supplementation decreased hepatic lipid peroxide and activated antioxidant enzymes levels in high-fat fed mice [15], and it has also been reported the UF demonstrated potent anti-inflammatory activity in an ovalbumin-induced mouse model of allergic airway inflammation [16]. However, the antiinflammatory and antioxidant properties of UF have not been investigated in the context of alcohol-induced liver damage. Therefore, we investigated the effect of UF on chronic alcoholinduced inflammation and on the antioxidant system in rats.
MATERIALS AND METHODS
Animals and diets
Twenty-four male Sprague-Dawley rats (4-week-old) were purchased from Orient Bio Inc. (Seoul, Korea). After a one week adaptation period, animals were randomly divided into a normal control group (NC), an alcohol control group (Al), or a UF (0.05 g/L in diet, Sigma, St. Louis, MO, USA) supplemented with alcohol diet group (Al-UF). Animals were placed on these diets for 8 weeks, and were individually housed in stainless-steel cages in an air-conditioned environment at 20 ± 2℃, and relative humidity 50 ± 5% under a 12-h light. The compositions of diets are shown in Table 1. The rats in the two alcohol groups were given a liquid alcohol diet (36% of energy) in which ethanol was introduced progressively. Specifically, animals were provided with 3% ethanol for the first 2 days (21% of energy), 4% for the next 2 days (28% of energy) and 5% (36% of energy) thereafter [17]. Normal control rats received an isocaloric liquid diet containing dextrin-maltose instead of ethanol. The rats in the Al and Al-UF groups received food ad libitum, whereas NC group received the same amount of diet that the alcohol control rats consumed the previous day. The study was approved by the Sunchon National University Institutional Animal Care and Use Committee (SCNU-IACUC-2012-7).
At the end of the 8-week experimental period, animals were anesthetized with CO2 gas after a 12-h fast. Blood was then drawn from inferior vena cava into tubes, and serum was obtained by centrifuging the blood at 900 ×g for 15 min at 4℃. The organs were then removed, rinsed with physiological saline, and immediately weighed. Serum and organ samples were stored at -70℃ until required for analysis.
Serum cytokines levels
Serum cytokines levels were measured using a multiplex detection kit (M60-009RDPD, Bio-Rad, Hercules, CA, USA). Capture antibodies directed against the cytokines, TNF-α, IL-6, interleukin 10 (IL-10), interleukin 1β (IL-1β) and interferon γ (IFNγ), were covalently coupled to beads, and then reacted with serum. After washing several times to remove unbound protein, a biotinylated detection antibody was added to create a sandwich complex. The final detection complex was formed by adding streptavidin-phycoerythrin conjugate. Phycoerythrin served as a fluorescent indicator or reporter. All samples were assayed in duplicate and analyzed using a Luminex 200 Labmap system (Luminex, Austin, TX, USA). Data analysis was performed using Bio-Plex Manager software version 4.1.1 (Bio-Rad, Hercules, CA, USA).
Genes expression of inflammation and antioxidant enzymes
Liver was homogenized in Trizol reagent (Invitrogen Life Technologies, Grand Island, NY, USA), and total RNA was then isolated according to the manufacturer's instructions. DNase was used to remove DNA contamination, and the RNA was then re-precipitated in ethanol to ensure the absence of phenol contamination. For quality control, RNA purity and integrity were evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, USA). Total RNA (1 µg) was reverse-transcribed into cDNA using a QuantiTect® reverse transcription kit (Qiagen, Hilden, Germany), and then mRNA expressions were quantified by real-time quantitative PCR using a SYBR green PCR kit (Qiagen, Hilden, Germany) and the CFX96TM real-time system (Bio-Rad, Hercules, CA, UAS). The sequences of the primers used are shown in Table 2. Cycle thresholds were determined using SYBR green emission intensities during the exponential phase. Fold changes were determined using the 2-ΔΔCt method [18]. GAPDH was used as the internal control.
Antioxidant enzyme activities
Superoxide dismutase (SOD) activity was measured spectro-photometrically using a method based on the inhibition of superoxide-mediated reduction [19]. One unit was defined to be the amount of enzyme that inhibited the oxidation of pyrogallol by 50%. Catalase (CAT) activity was measured by monitoring the disappearance of hydrogen peroxide spectro-photometrically at 240 nm for 5 min, as previously described [19]. Glutathione peroxidase (GSH-Px) activity was also measured spectrophotometrically using a reaction mixture containing 1 mM glutathione, 0.2 mM NADPH and 0.24 units of glutathione reductase in 0.1 M Tris-HCl (pH 7.2) buffer. The reaction was initiated by adding 0.25 mM H2O2 and absorbance was measured at 340 nm for 5 min [19]. Glutathione-S-transferase (GST) activity was evaluated, as described by Habig et al. [20] using 1-chloro-2,4-dinitrobenzen (CDNB) as substrate. For analysis, the absorbances were measured at 340 nm in 0.1 M potassium phosphate buffer (pH 6.5) containing 60 mM CDNB, and 10 mM GSH.
Levels of GSH, H2O2, and lipid peroxide
GSH level was measured as previously described by Ellman [21]. Mitochondrial hydrogen peroxide (H2O2) levels in liver were measured using Wolff's method [22], and results are expressed as micromoles of H2O2 per milligram of mitochondrial protein. Malondialdehyde (MDA) concentration in the liver (a marker of lipid peroxidation) was measured as described previously [19].
Hepatic histology
Hepatic tissue was removed from each rat and connective tissues were removed. Samples were then fixed in 10% (v/v) paraformaldehyde/PBS, embedded in paraffin, and sectioned at 3-5 µm, and stained with Masson's trichrome to visualize collagen fibers in connective tissues. Sections were viewed under a microscope at 200×.
RESULTS
Effects on serum cytokines levels
Chronic alcohol intake induced significant increases in serum TNF-α and IL-6 levels and decreased IL-10 level; however, these changes were prevented by UF supplementation. IL-1β and IFN γ levels did not differ between groups (Fig. 2).
Effects on the expressions of inflammatory genes
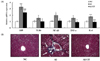
The mRNA expressions of LBP, TLR4, and NF-κB in the Al group were higher than in the NC group, but these changes were attenuated by UF supplementation. TNF-α and IL-6 mRNA expressions were up-regulated in the Al group as compared with the NC group; however, UF supplementation inhibited these up-regulations (Fig. 3A).
Effects on hepatic histology
Liver sections in the Al group revealed increased deposition of collagen fibers around congested central vein, indicating alcohol-induced fibrosis; however, collagen fiber deposition was less severe in the Al-UF group (Fig. 3B).
Effects on the genes expression and activities of antioxidant enzymes
The mRNA expressions of SOD, CAT, and GSH-Px were significantly down-regulated in the Al group as compared with the NC group, but UF up-regulated SOD and CAT mRNA levels compared to the Al group (Fig. 4). The activities of SOD, CAT, GSH-Px, and GST were also significantly lower in the Al group than in the NC group. However, SOD and CAT activities were significantly elevated by UF supplementation (Table 3).
Effects on levels of GSH, H2O2, and lipid peroxide
H2O2 levels were significantly lower in the Al-UF group than in the Al group, in which levels were similar to those in the NC group. Hepatic MDA levels in the Al group were 25% higher than in the NC group, whereas GSH levels were by 8% lower in the Al group. However, UF supplementation attenuated these changes (Table 3).
DISCUSSION
UF is a coumarin derivative with antioxidant and anti-inflammatory effects [23,24]. Mohamed et al. [25] found that UF ameliorated CCl4-induced oxidative stress by activating nuclear factor erythroid 2-related factor 2-mediated heme oxygenase-1 in rats. However, anti-inflammatory and antioxidant properties of UF have not been previously investigated in the context of alcohol-induced liver damage. The current study shows that LBP and TLR4 mRNA levels were significantly elevated in the liver of chronic alcohol-fed rats, but that these changes were attenuated by UF supplementation. LPS is a major factor in the pathogenesis of ALD and an inducer inflammation [6]. The recognition of this endotoxin is mainly initiated by LBP, a soluble serum lipid transfer protein synthesized by the liver, whose function is to extract LPS monomers from aggregated endotoxin structures for subsequent delivery to membrane-bound or soluble CD14 [26]. Thus, the hepatic expression of the LBP gene reflects LPS influx into liver. Furthermore, the activation of TLR4 by LBP leads to the rapid activation of NF-κB, and thus, to the production of several inflammation mediators, such as, TNF-α and IL-6 [27]. In the present study, NF-κB mRNA levels were reduced by UF, which indicates a reduction in inflammatory response, as was confirmed by significant decreases in hepatic expression of TNF-α mRNA.
Pro-inflammatory cytokines, such as, TNF-α, have recently been shown to play major roles in pathogenesis of liver disease, and their serum levels have been shown to be enhanced in ALD patients and in animal models of alcohol-induced liver damage [5]. TNF-α has been suggested to mediate the early stage of fatty liver disease and transition to more advanced disease [28]. Pro-inflammatory cytokines (IL-1β, TNF-α, IL-6 or IFNγ) and anti-inflammatory cytokines (IL-10) are produced in response to LPS [29,30]. In the present study, we also found that UF supplementation decreased serum TNF-α and IL-6 levels, but increased IL-10 levels in chronic alcohol-fed rats. IL-10 is one of the most important anti-inflammatory cytokines and has been associated with the amelioration of liver inflammation in different models [31]. In addition, IL-10 controls the productions of other cytokines, such as, IL-6 and TNF-α, decreases T-cell activation [32,33], and exerts hepatic protective effects against proliferation and fibrosis [34]. On the other hand, elevated hepatic and serum levels of IL-6 have been associated with the pathogenesis of alcoholic liver injury in animal models and ALD patients, in whom serum IL-6 levels were found to be positively correlated with disease severity [27,35]. In the present study, these augmenting effects of UF on the anti-inflammatory cytokine production and pro-inflammatory cytokine suppression strongly enhanced its anti-inflammatory effect in chronic alcohol-fed rats. Thus, our findings suggest UF could protect against alcohol-induced liver fibrosis by suppressing alcohol-induced TLR4-mediated hepatic fibrosis.
Excessive chronic exposure to alcohol usually reduces antioxidant defenses in hepatic tissue and blood [36], and thus, maintenance of the antioxidant system is essential in relation to the prevention of ALD [37]. Our results show that chronic alcohol feeding decreased the hepatic genes expression and activities of antioxidant enzymes (SOD, CAT and GSH-Px) and GSH levels. Previous studies have shown diminished SOD levels are associated with the risk of cell injury [38]. SOD may play an important role in protecting cells and tissues against the toxic effects of superoxide radicals [5]. CAT, a heme protein enzyme, has been reported to remove H2O2 [39], and thus, reductions in CAT activity might reflect reduced production and/or its inhibition as a result of the increased production of free radicals [40]. We found that, UF significantly increased the activities and mRNA levels of SOD and CAT, but not of GST and GSH-Px, as compared with the Al group, and that this was associated with reduced mitochondrial H2O2 and MDA levels. MDA is a secondary metabolite produced by lipid peroxidation in cell membranes, and the amount of MDA is used as an indicator of lipid peroxidation [41]. Interestingly, in the present study, we found that hepatic MDA levels were lower in the UF-supplemented alcoholic rats, which suggests the protection afforded by UF against alcohol-induced liver injury via enhancement of antioxidant defense system. In our previous study, UF supplementation was found to decrease serum ALT and γGTP activities effectively and to suppress body weight loss by alcohol intake without changing food intake or liver weight in chronic alcohol-fed rats [42].
In conclusion, the results of the present study indicate that UF protected against alcohol-induced liver damage by inhibiting the TLR4 signaling pathway and improving the antioxidant defense system. Therefore, we suggest UF be viewed as a promising therapeutic strategy for the treatment of alcoholic liver damage.




 XML Download
XML Download