

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Gliomas have a very high mortality rate owing to a peculiar feature of the tumor-rapid cell migration-that cannot be controlled either by surgery or radio-irradiation. Therefore, glioma patients generally only survive for an average of one year from the time of tumor development. Gliomas are not only difficult to diagnose but are also difficult to treat, primarily because their pathology and pattern of invasion are still poorly understood [1,2,3].
Cell invasion coupled with increased rates of cell proliferation, neoangiogenesis, and extensive infiltration of brain parenchyma are characteristic features of progression of brain cancer [4,5]. The mitogen-activated protein kinase (MAPK) cell-signaling pathway is made up of three major proteins: p38 MAPK, Jun N-terminal kinase, and extracellular signal-regulated kinase 1/2 (ERK1/2). These have been reported to be the active molecules involved in the control of cell proliferation and migration in a diverse range of cell types. [6]. Specifically, ERK1/2 has been implicated in cell migration and proliferation in most cancer cells [7]. Matrix metalloproteinases (MMP) are extracellular endopeptidases that have been associated with tumor progression owing to their involvement in the destruction of the basal membranes of cells [8]. Hence, activation of MMP9 leads to high motility and an increased invasion capacity in cancerous cells. Elevated expression of the tissue inhibitor of metalloproteinases-1 (TIMP-1) can inhibit the expression of MMP9 [9,10,11].
Samallanthus sonchifolius, commonly known as yacon, is a foodstuff that promotes human health. Recently, several studies have suggested that a number of important functional foods are potential candidates as medicinal plants and herbs in traditional Korean Medicine due to their pharmacological and biological properties. Yacon consists of diverse bioactive compounds that do not only induce physical health effects but also promote physiological responses. Yacon contains phytoalexins for antimicrobial activity, chlorogenic acids as antioxidant, anti-inflammation, anti-allergic, anti-cancer, and probiotic properties that counteract obesity and diabetes [12,13,14,18,19,20]. Although, it has been widely reported that yacon exhibits an anti-cancer effect, its therapeutic effect on gliomas remains to be elucidated. In the present study, we demonstrate that yacon extract inhibits cell migration by down-regulating the expression of MMP9 and decreasing the phosphorylation of ERK1/2. These findings provide a mechanistic insight into how yacon extract suppresses glioma cell growth and progression.
MATERIALS AND METHODS
Chemicals
Cell culture materials were purchased from Gibco BRL (Gaithersburg, USA). The EZ-Cytox Cell Viability Assay Kit was purchased from Daeil Lab Service (Seoul, Korea). GAPDH, ERK 1/2, and phospho-ERK1/2 antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, USA). All other chemicals were purchased from Sigma corporation.
Ethanol extraction of plant material
One-hundred grams of the plant were blended, and the crude powder was extracted in 1000 mL of sterile deionized water at 100℃ for 3 h. The aqueous extract was concentrated up to 300 mL at 60℃ under vacuum by the rotary evaporator. The remaining debris was precipitated with 700 mL absolute ethanol at 4℃ for 3 days. The supernatant was discarded and then was concentrated at 60℃ under vacuum by the rotary evaporator. The concentrated extract was dissolved in 50 mL of sterile deionized water and then lyophilized by freeze-drying at -60℃.
Cell culture and viability assay
Rat C6 glioma cells were obtained from the Korean Cell Line Bank (Seoul, Korea) and were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin at 37℃ in a 5% CO2 atmosphere. C6 glioma cells were seeded at 5 × 104 cells/mL in a 96-well microplate containing DMEM and were then incubated for 24 h. The cells were incubated with different concentrations of yacon extract (100, 200, and 300 µg/mL) in either FBS or FBS-deficient media for 24 h. Cell viability was determined using an EZ-Cytox Cell Viability Assay Kit. The cell viability was determined as a percentage by comparing the absorbance at 450 nm for the treated cells and untreated control cells that had been incubated in FBS-deficient media.
Scratch wound healing assay
C6 glioma cells were seeded at a density of 1 × 105 cells/mL in six-well plates and incubated in an FBS-containing medium for 24 h. The medium was then replaced with a serum-free medium. Following serum starvation for 24 h, a scratch wound was made across the center of the monolayer of cells in each well by using a sterile 200-µL pipette tip. This was followed by incubation with or without yacon extract in a serum-containing medium for an additional 24 h. Images of the cells that had migrated into the cell-free scratch wound area were acquired using an inverted microscope (IX71; Olympus, Tokyo, Japan) fitted with a charge-coupled device (CCD) camera. The migration distance was measured using the ImageJ software package. The scratch wound areas were determined by the relative percentage compared to untreated control cells in FBS-deficient media.
Boyden chamber assay
Boyden chamber assay was performed as previously reported [15]. To determine the effect of yacon extract on the migration of C6 glioma cells, the Boyden chamber assay was performed in a 48-well chemotaxis chamber (Neuro-Probe, Gaithersburg, MD, USA). Briefly, polycarbonate filter membranes (Neuro-Probe) with a pore size of 8 µm were coated with 0.1% collagen type-I and dried at room temperature for 1 h. Two different concentrations of yacon extract (200 and 300 µg/mL in 10% FBS media) were prepared and then 27.8-µL portions were dispensed into the lower chamber of the wells. The lower chamber containing the extract was then covered by the coated membrane, and 50 µL of C6 glioma cells (in serum-free media) were loaded into the upper chamber at a concentration of 1 × 106 cells/mL. Following culture at 37℃ in 5% CO2, cells on the lower surface of the membrane were fixed with methanol and stained using Diff-Quick. After staining, cells that had migrated through the filter membrane were imaged and counted using an inverted microscope as described above.
Western blot analysis
To analyze protein expression, we performed western blotting using specific antibodies. Briefly, 20 µg of protein was prepared for each treatment group. After boiling, the proteins were separated by electrophoresis on 12% acrylamide gels and then transferred onto polyvinylidene difluoride membranes in transfer buffer at 4℃ for 2 h. The membrane was blocked with 5% BSA in Tris-buffered saline (TBS) at room temperature for 1 h and then washed in TBS with 0.1% Tween 20 (TBS/T). The membranes were incubated overnight at 4℃ with either phosphorylated ERK1/2 (p-ERK1/2) antibody, or ERK1/2 antibody, both at a 1 : 2000 dilution. The membranes were washed with TBS/T, followed by incubation with a 1 : 5000 dilution of IgG secondary antibody conjugated to horseradish peroxidase. The expression levels of the proteins were analyzed using chemiluminescence (ECL plus kit; Amersham Pharmacia Biotech). Developed protein bands were visualized and quantified using Image J Software.
Reverse transcription PCR
Total RNA was isolated from cells using TRI-reagent and then cDNA was synthesized using 0.5 µg of total RNA; the superscript II reverse transcription system with oligo-deoxythymidine. PCR amplification was performed using the following protocol: a predenaturation at 95℃ for 3 min and then either 35 cycles of denaturation at 94℃ for 35 s, annealing at 52℃ for 35 s, and extension at 72℃ for 35 s, followed by a final extension at 72℃ for 10 min. The mRNA expression was quantified using an ethidium bromide-stained 1.5% agarose gel. The following primers were used for RT-PCR: MMP9, sense primer, 5'-AAGTTTGGTGTCGCGGAGCAC-3', and anti-sense primer, 5'-TACATGAGCGCTTCCGGCAC-3'; TIMP-1, sense primer, 5'-ACAGCTTTCTGCAACTCG-3', and anti-sense primer, 5'-CTATAGGTCTTTACGAAGGCC-3'; GAPDH, sense primer, 5'-GGCATTGCTCCTCAATGACAA-3', and anti-sense primer, 5'-TGTGAGGGAGATGCTCAGTG-3'.
Statistical analysis
The results are expressed as the mean ± standard error (SE) of at least three independent experiments. The difference between the two groups was examined using Student's t-test, and one-way analysis of variance (ANOVA) with Tukey's test was used for multiple comparisons (Prism 4.00 for Windows, GraphPad, San Diego, USA). A P-value of < 0.05 was considered as statistically significant.
RESULTS
The effect of yacon extract on the viability of FBS-stimulated C6 glioma cells
To determine the effect of yacon extract on the viability of FBS-stimulated C6 glioma cells, we performed a XTT cell viability assay. To determine the effect of yacon extract on the FBS-stimulated cell growth of glioma cells, C6 glioma cells were treated with yacon extract (100, 200 and 300 µg/mL) in either FBS-deficient media or in 10% FBS for 24 h. As shown in Fig. 1, the cell viability was unaffected by yacon extract up to a concentration of 300 µg/ml. The percentage cell growth was 146 ± 2.6% in cells stimulated by 10% FBS. The FBS-induced C6 cell growth was reduced by 40.7 ± 4.1% in response to 300 µg/mL of yacon extract.
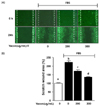
The effect of yacon extract on scratch wound healing in FBS-stimulated C6 glioma cells.
The scratch wound healing assay is commonly used as a method to test the migration rate for a variety of cell types. In this study, we carried out the scratch wound healing assay, to test the effect of yacon extract on the cell migration rate of FBS-stimulated C6 glioma cells. As shown in Fig. 2, the results showed a significant increase in cell motility of up to 222.4 ± 10% when cells were post-treated with 10% FBS in comparison to untreated cells. On the other hand, the FBS-stimulated C6 cells exhibited a reduction of 50.8 ± 15.8% and 88.5 ± 12.4% in the migration rate when treated with yacon extract at concentrations of 200 µg/mL and 300 µg/mL, respectively.
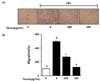
Yacon extract inhibits cell migration in FBS-stimulated C6 glioma cells
We performed the Boyden chamber assay to confirm the inhibitory effect of yacon extract on cell migration in FBS-stimulated C6 glioma cells. As shown in Fig. 3, the presence of 10% FBS significantly increased cell migration by 393.1 ± 12.3%. Treatment with 200 µg/mL and 300 µg/mL yacon extract reduced FBS-induced C6 cell migration by 222.0 ± 8.3% and 372.5 ± 12.7%, respectively.
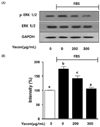
Yacon extract suppresses the phosphorylation of ERK 1/2 in FBS-stimulated C6 glioma cells
Western blot assays were performed to analyze the degree of phosphorylation of ERK1/2 in FBS-stimulated C6 glioma cells. As shown in Fig. 4, the expression of the phosphorylated ERK1/2 was increased by 75.33 ± 6.8% in cells stimulated by 10% FBS, compared to those not treated with FBS or exposed to yacon extract. In addition, yacon extract significantly reduced the phosphorylation of ERK1/2 by 33.0 ± 1.7% and 69.52 ± 5.2% at concentrations of 200 µg/mL and 300 µg/mL of yacon extract respectively.
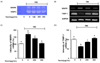
Yacon extract suppressed MMP9 mRNA expression by inducing TIMP-1 expression in FBS-stimulated C6 glioma cells
Cell migration plays an important role in glioma cell progression, a phenomenon that is associated with increased MMP9. In contrast, the MMP9 activity can be controlled by the TIMP-1. Therefore, we employed reverse transcription PCR using specific primers to determine whether yacon extract could affect the mRNA expression levels of MMP9 and TIMP-1 in FBS-stimulated C6 glioma cells. As shown in Fig. 5, the elevation of MMP9 expression was enhanced by 10% FBS resulting in an increase of 75.15 ± 5.0%. Yacon extract significantly reduced the expression of MMP9 by 32.0 ± 4.9% and 69.66 ± 4.7% at concentrations of 200 µg/mL and 300 µg/mL of yacon extract respectively. FBS (10%) reduced TIMP-1 expression by 37.97 ± 4.1%. In contrast, yacon extract increased the expression of TIMP-1 by 23.67 ± 3.5% and 44.33 ± 3.9% at concentrations of 200 µg/mL and 300 µg/mL, respectively.
DISCUSSION
Samallanthus sonchifolius otherwise known as yacon, is widely cultivated worldwide and has been found to contain numerous carbohydrate and secondary metabolites. Several recent studies have demonstrated the medical benefits of yacon, which are related to its effects on various cellular signaling molecules [12,13,14]. However, none of these studies has yet established the effect of yacon on brain tumors. To address this we have attempted to investigate the components of yacon and its effects in preventing the progression of glioma. This study suggests that yacon contains components that can prevent glioma progression.
Glioma is associated with aggressive tumor cell motility that contributes to the rapid metastasis of the cancer cells in to normal tissues. This leads to a lack of a distinct boundary between normal tissues and the tumor, limiting the possibility for effective surgery [2]. Martin et. al. have suggested that antioxidants may control glioma cell proliferation [19]. Campos et al. have shown that yacon has a high antioxidant content [20]. Although cancer progression is associated with complex signaling pathways, normally the phenomenon of metastasis involves migration of cells [21]. We explored the inhibitory effect of yacon on the migration of C6 glioma cells using noncytotoxic concentrations of yacon. Our data from the scratch wound healing assay and the Boyden chamber assay showed that yacon inhibited abnormal cell motility in FBS-induced C6 glioma cells in a dose-dependent manner.
The invasion and migration of malignant cells play an important role in the progression of cancer from one stage to another [22]. Recent advances in cancer therapy suggest that controlling cell migration by targeting molecular function is a promising treatment strategy [23]. ERK1/2 contributes to cell migration and proliferation in a number of different types of cancer cell [6]. Recent extensive analysis of fetal bovine serum (FBS) has demonstrated that it contains complex components not present in newborn calf serum (NCS). These active components are believed to directly influence the proliferation and differentiation of cells [24], as well as activate signaling pathways, such as MAPK signaling pathway, involved in cell migration and invasion [25]. A number of studies have shown that specific growth factors directly influence cell mobility patterns found in a number of different cancers including lung cancer and glioma [26,27,28]. Antioxidant proteins, which are activated via tyrosine kinase receptor (RTK) pathways, are known to counter-regulate abnormal cell growth in tumor cells. For instance, N-acetylcysteine (NAC) has been shown to regulate the proliferation of glioma cells by regulating cell cycle arrest at the G0/G1 transition and by up-regulating expression of p21 [16]. In this study, we examined the effects of yacon on C6 glioma cells and tested whether it could suppress the phosphorylation of ERK1/2 a component of the MAPK pathway. Our results show that yacon significantly decreased the phosphorylation of ERK1/2 in FBS-stimulated C6 glioma cells. The major finding of our study is that the suppression of cell migration in FBS-stimulated C6 glioma cells by yacon is mediated by the ERK1/2 pathway. Therefore, these results suggest that yacon is a potential functional food candidate that can act as an antioxidant and is therefore potentially useful in preventing glioma progression.
Furthermore, we found that the expression of MMP9 was significantly inhibited by yacon in a concentration-dependent manner. MMP9 interacts with major intracellular signal transduction pathways causing the extracellular matrix (ECM) destruction and is a well-known regulator of the cancer cell migration pathway [10]. For these reasons, we investigated the concentration of yacon required to inhibit or reduce the expression of MMP9. We also tested whether yacon could induce TIMP-1 expression. Interestingly, the expression levels of TIMP-1 were also reduced by yacon in a concentration-dependent manner.
In conclusion, we suggest that yacon is a potential candidate for the treatment of malignant gliomas because of its ability to significantly reduce the motility of C6 glioma cells. This effect was found to occur through modulation of the phosphorylation of ERK1/2 and regulation of MMP9/TIMP-1 signaling. The major finding of our study is that yacon contributes to the suppression of cell migration of FBS-stimulated C6 glioma through inhibition of the ERK1/2 and MMP9/TIMP-1 signaling pathways. However, based on the present data, we cannot specify the exact amount of yacon intake required to prevent the migration of glioma in the brain. Because our data was obtained in vitro, further in vivo experiments are required to confirm these results.

 XML Download
XML Download