

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Inflammation plays a pivotal role in various diseases, such as cardiovascular disease, cancer, rheumatoid arthritis, asthma and bronchitis [1,2]. Among inflammation-related processes, macrophages play an important role in inflammatory diseases and secrete pro-inflammatory cytokines including interleukin-1 beta (IL-1β), IL-6 and tumor necrosis factor-alpha (TNF-α) and inflammation mediators including nitric oxide (NO) and prostaglandins [1]. In addition, inflammatory enzymes such as inducible nitric oxide synthase (iNOS) and cyclooxyenase-2 (COX-2) are associated with development of numerous inflammatory diseases [3]. Thus, inhibition of production of inflammatory mediator is the major therapeutic target for treatment of inflammatory diseases.
Zebrafish has become a useful model system for vertebrate development because of its large clutches, small size, transparency, low cost and physiological similarity to mammals [4,5]. In addition, zebrafish has an advantage for in vivo study because of the rapid development and optical transparency during development [4,7]. Recently, zebrafish has emerged as a model system for use in pharmacological studies for human disease such as inflammation, angiogenesis, hypolipidemic and skin aging [5,6,7,8].
Styela clava, used in this study, is an important cultured aquatic species used for food by people in Korea. Several studies on S. clava have pointed out a variety of biological benefits, including antioxidative [9], anticancer [10] hepatoprotective [11] and antihypertensive effects [12,13]. Although the protein content of S. clava flesh tissue is more than 60%, there remains a paucity of data regarding the biological activities of S. clava flesh tissue protein.
Controlled enzymatic hydrolysis of food proteins may also modify and improve and upgrade the nutritional and functional properties of food proteins [14,15]. The molecular weight of the hydrolyzed food protein is a pivotal factors for effective production of protein hydrolysates to use as functional materials [15]. An ultrafiltration (UF) membrane system is used to obtain various molecular weight fractions of hydrolysates [15,16]. The UF system has the major advantages that the molecular weight distribution of hydrolysates can be controlled by appropriate selection of the UF membrane [17].
The current study was conducted in order to investigate the potential effect of enzymatic hydrolysates and molecular weight fractions from S. clava flesh tissue on inflammation through an in vitro study by determining the effect on LPS-induced into macrophages as well as in an in vivo zebrafish model.
MATERIALS AND METHODS
Materials
Styela clava was collected between March 2010 and May 2011, and kindly donated by Miduduk Corporated Association (Masan, Korea). Commercial food grade proteases including Protamex, Kojizyme, Neutrase, Flavourzyme and Alcalase from Novo Co. (Novozyme Nordisk, Bagasvaerd, Denmark). Dublecco's modified eagle's medium (DMEM), fetal bovine serum (FBS) and phosphate buffer saline (PBS) were purchased from Gibco BRL Co. (Gaithersburg, MD, USA). Dimethyl sulfoxide (DMSO), 2',7'-Dichlorodihydrofluorescein diacetate (DCFH-DA), and 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazoliumbromide (MTT) were purchased from Sigma Co. (St. Louis, MO, USA). The other chemicals and reagents used in this study were of experimental grade.
Preparation of S. clava flesh tissue protein hydrolysates (SFTPH) and molecular weight fractions
The hydrolytic enzymes used in preparation of enzymatic hydrolysates, eight proteases (Protamex, Kojizyme, Neutrase, Flavourzyme, Alcalase, pepsin, tyrpsin and papain) hydrolyzed the powder of S. clava flesh tissue. The dried ground S. clava flesh tissue powder was homogenized with buffer, and hydrolyzed with enzymes in a substrate to enzyme ratio of 100 : 1 (w/w). The pH of the homogenates was adjusted to its optimal pH value before the enzymatic hydrolysis. The enzymatic reactions were performed for 12 h in order to achieve an optimal degree of enzymatic hydrolysis (Table 1). The hydrolysates were then boiled for 10 min at 100℃ in a water bath to inactivate the enzyme reaction. Each enzymatic hydrolysate was clarified by centrifugation (3500 rpm, for 20 min at 4℃) for removal of the residue. The resultant SFTPH was fractionated through UF membranes with a range of molecular weight (MW) cut-off of 5 and 10 kDa, respectively. Fractionates were designed as follows: SFTPH-I with distribution of above 10 kDa, SFTPH-II with distribution of 5~10 kDa and SFTPH-III with distribution of below 5 kDa. All recovered SFTPH fractions were lyophilized and stored under -20℃ until used.
Cell culture
The murine macrophage cell line RAW 264.7 was purchased from the Korean macrophages Line Bank (KCLB; Seoul, Korea). RAW 264.7 macrophages were grown in DMEM (GIBCO INC., NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; GIBCO), streptomycin (100 mg/ml), and penicillin (100 unit/ml). Cultures were maintained at 37℃ in 5% CO2 incubator.
Cytotoxicity assay using MTT assay
RAW 264.7 macrophages (1.0 × 105 cells/ml) plated in a 96-well plate were pre-incubated and then treated with lipopolysaccharide (LPS; 1 µg/ml) plus aliquots of the hydrolysates in sterile distilled water. The cells were then incubated for an additional 24 h at 37℃. MTT stock solution (50 µl; 2 mg/ml in PBS) was then added to each well to a total reaction volume of 250 µl. After incubation for 4 h, the plates were centrifuged (800 × g, 5 min), and the supernatants were aspirated. The formazan crystals in each well were dissolved in 150 µl of dimethylsulfoxide (DMSO), and the absorbance was measured using an ELISA plate reader at 540 nm.
Determination of nitric oxide (NO) production
After pre-incubation of RAW 264.7 macrophages (1.0 × 105 cells/ml) with LPS (1 µg/ml) for 24 h, the quantity of nitrite accumulated in the culture medium was measured as an indicator of NO production. In brief, 100 µl of cell culture medium was mixed with 100 µl of Griess reagent [1% sulfanilamide and 0.1% naphthylethylenediamine dihydrochloride in 2.5% phosphoric acid], the mixture was incubated at room temperature for 10 min, and the absorbance at 540 nm was measured in a microplate reader. Fresh culture medium was used as a blank in every experiment.
Determination of PGE2 production
The three molecular weight fractions (SFTPH-I, SFTPH-II and SFTPH-III) were investigated for NO inhibitory activity. Among the three fractions, the highest NO inhibitory fraction (active fraction) was determined for cytokine inhibitory activities. The active fraction was diluted with DMEM prior to treatment. Cells were treated with LPS (1 µg/ml) for 24 h to allow cytokine production. The PGE2 concentration in the culture medium was quantified using a competitive enzyme immunoassay kit (R&D Systems, Minneapolis, MN, USA) in accordance with the manufacturer's instructions. Production of PGE2 was measured relative to that observed after control treatment.
Measurement of pro-inflammatory cytokine (TNF-α, IL-β and IL-6) production
The active fraction was solubilized with sterile distilled water before treatment. The inhibitory effect of the active fraction on pro-inflammatory cytokine (TNF-α, IL-β and IL-6) production from LPS (1 µg/ml) treated RAW 264.7 macrophages was determined as described by Cho et al. [18]. Supernatants were used for pro-inflammatory cytokine assay using a mouse ELISA kit (R&D Systems, Minneapolis, MN, USA).
Western blot analysis
The cell lysates were prepared with lysis buffer (20 mM Tris, 5 mM EDTA, 10 mM Na4P2O7, 100 mM NaF, 2 mM Na3VO4, 1% NP-40, 10 mg/ml aprotinin, 10 mg/ml leupeptin and 1 mM PMSF) for 60 min and then centrifuged at 12,000 rpm for 15 min at 4℃. The protein concentrations were determined using a BCA™ protein assay kit. The lysates containing 30 µg of protein were subjected to electrophoresis on sodium dodecyl sulfate (SDS)-polyacrylamide gel, and the gel was transferred onto a nitrocellulose membrane. The membrane was blocked in 5% non fat dry milk in TBST (25 mM Tris-HCl, 137 mM NaCl, 2.65 mM KCl, 0.05% Tween 20, pH 7.4) for 2 h. The primary antibodies were used at a 1 : 1000 dilution. Membranes were incubated with the primary antibodies at 4℃ overnight. The membranes were then washed with TBST, followed by incubation with the secondary antibodies used at 1 : 3000 dilutions. Signals were developed using an ECL western blotting detection kit and exposed to a Western imaging system.
Origin and maintenance of parental zebrafish
All animal procedures were performed in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Utilisation Committee for Veterinary Medicine of Jeju National University. Adult zebrafishes were purchased from a commercial dealer (Seoul aquarium, Korea) and 10 fishes were kept in a 3 L acrylic tank with the following conditions; 28.5℃, with a 14/10 h light/dark cycle. Zebrafishes were fed three times a day, 6 d/week, with tetamin flake food supplemented with live brine shrimps (Artemia salina). Embryos were obtained from natural spawning, which was induced in the morning by turning on the light. Collection of embryo was completed within 30 min.
Waterborne exposure of embryos to active fraction
From approximately two days post-fertilization (dpf), embryos (n = 25) were transferred to individual wells of a 24-well plate and maintained in embryo media containing 1 ml of vehicle (sterile distilled water) or 50, 100 and 200 µg/ml of active fraction for 1 h followed by treatment with LPS (3 µg/ml) or co-treatment with active fraction and LPS.
Image analysis of intracellular NO generation
Image analysis of intracellular NO generation was performed according to the method used by Wigesinghe et al. [19]. Generation of NO in an inflammatory zebrafish model was analyzed using a fluorescent probe dye, diaminofluorophore 4-amino-5-methylamino-2-,7-difluorofluorescein diacetate (DAF-FM DA). Transformation of DAF-FM DA by NO in the presence of dioxygen generates highly fluorescent triazole derivatives. The embryos were treated with 50, 100 and 200 µg/ml of active fraction, and, 1 h later, LPS (3 µg/ml) was applied to the plate. Following active fraction and LPS treatment, the zebrafish embryos were transferred into 96-well plates, treated with DAF-FM DA solution (20 µg/ml), and incubated for 1 h in the dark at 28.5℃. After incubation, the zebrafish embryos were rinsed in fresh zebrafish embryo medium and anesthetized in tricaine methanesulfonate solution (1 : 500 dilutions) before observation. Fluorescence intensity of individual zebrafish larvae and embryos was quantified at an excitation wavelength of 485 nm and an emission wavelength of 535 nm, using a Perkin-Elmer LS-5B spectrofluorometer. Images of stained embryos were observed using a florescence microscope equipped with a Moticam color digital camera (Olympus Optical, JP/BX60).
RESULTS
Effect of enzymatic hydrolysates on cell viability
Prior to evaluation of the NO inhibitory effect of enzymatic hydrolysates, we first examined its cytotoxic effect on the viability of RAW 264.7 macrophages. Cytotoxicity of enzymatic hydrolysates was evaluated using the MTT assay at a concentration of 200 µg/ml. None of the enzymatic hydrolysates affected the cytotoxicity of RAW 264.7 macrophages (Fig. 1). Thus, those concentrations were used in subsequent experiments.
Effect of fractionation of enzymatic hydrolysates from S. clava on LPS-induced NO production
To evaluate the question of whether the enzymatic hydrolysates possess a potential anti-inflammatory effect in LPS-stimulated RAW 264.7 macrophages, we investigated inhibitory effect of nine hydrolysates on NO production (Fig. 1). Among the hydrolysates, Protamex hydrolysate exhibited the highest level of inhibitory effect on NO production. Thus, Protamex hydrolysate was selected for further experiments and fractionation of the hydrolysate using UF membranes is described. In this study, using an UF system, Protamex hydrolysate was fractionated into three individual fractions with three molecular weight (MW) groups of SFTPH-I (MW > 10 kDa), SFTPH-II (MW = 5~10 kDa), and SFTPH-III (MW< 5 kDa), using UF membranes (MW cut-off of 5 and 10 kDa. In the NO inhibitory activity assay (Fig. 2), the highest level of inhibitory effect on NO was exhibited by SFTPH-I.
Effect of SFTPH-I on LPS-induced PGE2 production and iNOS and COX-2 expression
The inhibitory effect of SFTPH-I on PGE2 production in LPS-induced RAW 264.7 macrophages showed a similar pattern to its effect on NO inhibition, in that SFTPH-I inhibited LPS-induced production of PGE2 (Fig. 3A). Although SFTPH-I had an inhibitory effect on PGE2 production, the effect was not as strong as that exhibited in the inhibition of NO production. To elucidate the mechanism involved in the inhibition of NO generation by SFTPH-I in LPS-induced RAW 264.7 macrophages, we further studied the effect of SFTPH-I on iNOS and COX-2 protein expression by Western Blot. In response to LPS expression of iNOS was markedly increased, and SFTPH-I inhibited iNOS and COX-2 protein (Fig. 3B). These results suggested that SFTPH-I inhibited release of NO and PGE2 by reducing expression of iNOS and COX-2.
Effect of SFTPH-I on LPS-induced IL-β, IL-6 and TNF-α production
To determine the effects of SFTPH-I on production of proinflammatory cytokines such as IL-1β, IL-6, and TNF-α, RAW 264.7 macrophages were incubated with SFTPH-I (50, 100 and 200 µg/ml) in the presence or absence of LPS (1 µg/ml) for 24 h, and the cytokine levels were measured by ELISA. Results showed that pretreatment of macrophages with SFTPH-I resulted in considerably reduced production of IL-1β, IL-6, and TNF-α (Fig. 4A-C).
Effect of SFTPH-I on LPS-induced MAPKs activation
To determine whether SFTPH-I inhibited the production of pro-inflammatory mediators through effects on the MAPK pathway, we evaluated the effect of SFTPH-I on LPS-induced phosphorylation of p38, JNK, and ERK in RAW 264.7 macrophages using western blotting. The levels of phosphorylated ERK, JNK, and p38 MAPK were elevated in macrophages after treatment with LPS only, whereas the levels of total ERK, JNK, and p38 MAPK did not show significant change (Fig. 5). However, compared to cells treated with LPS only, macrophages pretreated with SFTPH-I showed lower LPS-induced activation of these proteins (Fig. 5).
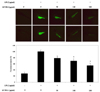
Inhibitory effect of NO generation by LPS in zebrafish
Prior to evaluation of the NO inhibitory effect of SFTH-I, we first examined its toxicity on the survival rate of zebrafish. SFTH-I and LPS did not affect the toxicity of zebrafish (Fig. 6). Fig. 7 shows a typical fluorescence micrograph of the zebrafish. We found that NO produced by LPS exposure in the yolk sac of zebrafish embryos. The negative control, which contained no SFTH-I or LPS, generated a clear image, whereas the positive control, which was treated with LPS, generated a fluorescence image, which suggests that NO production occurred during LPS treatment in the zebrafish. However, in zebrafish treated with SFTH-I prior to LPS treatment, a reduction in the amount of NO was observed.
DISCUSSION
NO is a pivotal inflammatory mediator synthesized from arginine by nitric oxide synthesis (NOS) [1,20]. Under pathological condition, NO is increased by the iNOS, which subsequently leads to tissue damage [21]. Thus, NO inhibitors are essential for prevention of inflammatory responses. Recent studies have investigated the effect of marine-derived hydrolysates on LPS-stimulated macrophages for treatment of inflammation [22,23]. In the present study, we examined the anti-inflammatory effect of enzymatic hydrolysates from S. clava flesh tissue on LPS-stimulated macrophages and an in vivo zebrafish model.
To obtain the active anti-inflammation hydrolysate, nine proteases were used in hydrolysis of S. clava flesh tissue protein. Protamex-proteolytic hydrolysate showed the highest NO inhibitory activity in macrophages and was selected from further fractionation using UF membrane. Recently, many studies have reported findings that enzymes are capable of producing bioactive properties when incorporated for hydrolysis of food proteins [24,25,26,27]. The molecular weight of the hydrolyzed food protein is a pivotal factor in production of protein hydrolysates with desired biological materials [17]. One of the methods for fractionation of various molecular weights of hydrolysates is to use UF membranes with different pore size [15,17]. Thus, Protamex hydrolysate was fractionated using an UF system into three individual fractions, and then NO inhibitory activity was examined in macrophages. NO inhibitory activity was widely observed in all of the fractions, thus suggesting that many NO inhibitory substances with various molecular weight ranges were contained in the hydrolysate. Among the fractions, the most potent NO inhibition was noted in an over 10 kDa fraction (SFTPH-I). Further study sought to elucidate the pharmacological effects of SFTPH-I on production of pro-inflammatory mediators, and its mechanism of action, in LPS-stimulated RAW 264.7 macrophages. NO is generated from L-arginine by NOS, a three member enzyme family, including iNOS. In addition, COX-2 is an important mediator of inflammation involving NO and PGE2 generation [28]. PGE2 has been implicated as an important mediator in the processes of inflammation [29]. As previously reported, generation of PGE2 is closely related to NO production [1]. Thus, chemopreventive agents that attenuate COX-2-mediated PGE2 production have had a therapeutic effect in many inflammatory diseases [30]. Our results demonstrated that SFTPH-I inhibited NO and PGE2 production in an LPS-stimulated RAW 264.7 macrophage culture system by reducing the expression of iNOS and COX-2.
In addition, SFTPH-I also reduced the LPS-induced production of TNF-α, IL-6, and IL-1β in a dose-dependent manner. TNF-α is a potent activator of macrophages and can stimulate the production or expression of IL-6, IL-1β, PGE2, collagenase, and adhesion molecules. It elicits a number of physiological effects, including septic shock, inflammation, and cytotoxicity [31]. IL-6, a well-known pro-inflammatory cytokine, is regarded as an endogenous mediator of LPS-induced fever [32]. IL-1β is regarded as a pivotal pro-inflammatory cytokine, primarily released by macrophages, and it plays an important role in the pathophysiology of rheumatoid arthritis [33]. Inflammatory stimuli, such as LPS, induce cytokines in the process of macrophage activation, which mediates tissue response in different phases of inflammation [34,35]. Because of their important roles in inflammatory responses, down-regulation of these pro-inflammatory cytokines is of utmost importance during anti-inflammatory therapy. Here, we showed that SFTPH-I inhibited the LPS-induced production of TNF-α, IL-1β, and IL-6.
MAPKs, including ERK, JNK, and p38 kinase play important roles in many biological processes, including inflammation, proliferation, and differentiation [31]. According to a previous report, MAPKs promote iNOS and COX-2 expression in LPS-stimulated macrophages [36]. In addition, phosphorylation of MAPKs promotes generation of pro-inflammatory cytokines in LPS-stimulated macrophages [37]. Thus, the inhibition of MAPKs activation or function is a key mechanism. SFTPH-I attenuated LPS-induced phosphorylation of ERK, p38 and JNK. Taken together, these results suggest the potential role of ERK, p38 and JNK in SFTPH-I-induced suppression of NO, and pro-inflammatory cytokines in activated macrophages.
The number of chemicals that need to be tested in the field of chemical toxicity and drug discovery is steadily increasing. Thus, the need for high-throughput screening methods also arises, where the use of zebrafish embryos was proposed [38] because of their small size and thus suitability for studies in multi-well plates. Not only toxicity screening applications are imaginable but also applications for clarification of mechanisms of toxicity have been studied [39]. In the current study, the potential NO inhibitory effect of SFTH-I was investigated in zebrafish as an alternative animal model system. When zebrafish were treated with SFTH-I prior to LPS treatment, a dramatic decrease in NO production was observed.
Recently, some anti-inflammatory properties have also been reported in certain bioresource proteins, including microalgae, salmon pectoral fin and mussel [40,41,42]. Potent anti-inflammatory peptides can be derived by enzymatic proteolysis of proteins and may act as potential physiological modulators of metabolism during gastrointestinal digestion of nutrients [43,44]. Bioavailability of nutritional supplements generally designates the proportion of the administered substance capable of being absorbed [45]. In terms of bioactive peptides, the bioavailable fractions are mainly short peptides that survive gastrointestinal digestion and absorption [46]. Therefore, further studies are needed for identification of the mechanism of enzymatic hydrolysate from S. clava flesh tissue for the potent anti-inflammatory effect in animal or human.
In conclusion, we evaluated the NO inhibitory effect of the enzymatic hydrolysates of S. clava protein. The Protamex hydrolysate was found to have the highest NO inhibitory effect and was fractionated using the UF membrane system. The above 10 kDa fraction (SFTPH-I) was a potent inhibitor of NO, PGE2, iNOS, COX-2, IL-6, IL-1β, and TNF-α production in LPS-induced RAW 264.7 macrophages. In addition, the levels of phosphorylated MAPKs in LPS-induced RAW 264.7 macrophages were decreased by treatment with SFTPH-I. SFTPH-I also demonstrated NO inhibitory properties against an LPS-stimulated zebrafish model. Based on the results of this study, it appears that this hydrolysate and molecular weight fraction may have a beneficial effect on treatment of inflammatory diseases.







 XML Download
XML Download