

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Oxidative stress and inflammatory process are mainly attributed to neurodegenerative diseases, including dementia, which has recently attracted significant attention. As brain cells die, interaction between neurons would be inhibited, resulting in impairment of memory and cognitive function, known as dementia [1,2]. Overproduction of free radicals leads to oxidative stress, which induces cells to go under an inflammation mechanism and lowers cell viability. In particular, nitric oxide (NO) reacts with membrane lipids to induce lipid peroxidation [3]. NO and superoxide (O2-) combine to form a highly reactive intermediate, peroxynitrite (ONOO-). ONOO- induces DNA strand breaks, lipid peroxidation, and protein nitration. By overproduction of NO and generation of ONOO-, glial cells would be damaged [4,5]. Injury of glial cells produces a variety of pro-inflammatory and neurotoxic factors, including nuclear factor kappa B (NF-κB), inducible nitric oxide synthases (iNOS), cycloxygenase-2 (COX-2), and several cytokines [3]. As above, this process can surely occur in C6 glial cells, in which the oxidative damage can lead to dementia. Therefore, attenuation of oxidative stress and inhibition of inflammatory process in glial cells are crucial for prevention of neurodegenerative diseases.
Natural extract products have recently attracted significant attention for prevention and treatment of degenerative diseases. Litchi chinensis Sonn., a subtropical fruit, is widely cultivated in Africa, South-East Asia, such as China, Taiwan, Vietnam, and Japan etc. [6,7]. The fresh fruit has a sweet flavor and contains sugars, citric acid, vitamin C, and polyphenols [8]. It has traditionally been used as a tonic for the function of heart and liver and it has been reported to exhibit strong antioxidant activities [9,10]. It contains polyphenolic compounds such as condensed tannins, epicatechin, procyanidin A2, anthocyanin, quercetin 3-rutinoside (rutin), and quercetin glucoside [10,11]. Oligonol is a phenolic product derived from Litchi chinensis Sonn. extract containing catechin-type monomers and oligomers of proanthocyanidins [12]. Emerging evidence indicates that oligonol has some biological effects [13], including anticancer, antioxidant, and anti-inflammatory effects, as well as beneficial activity for NO bioavailability and regulatory effect on lipid metabolism. However, study on the protective role of oligonol against oxidative stress in C6 glial cells has not yet been conducted. In the current study, we investigated the protective effect and its related mechanisms of oligonol against oxidative stress-induced inflammation in C6 glial cells.
MATERIALS AND METHODS
Oligonol
Oligonol was provided by Amino Up Chemical Co., Ltd. (Sapporo, Japan). Oligonol was produced from lychee fruit (Litchi chinensis Sonn.) extract using a patented technology process (international patent WO 2004/103988 AI) at Amino Up Chemical Co., Ltd. (Sapporo, Japan) [5]. Briefly, dried Litchi chinensis Sonn. was extracted with 50% [volume to volume (v/v)] ethanol. The filtrate was evaporated and passed through a DIAION HP-20 column, and eluted with ethanol. The eluate was then evaporated to dryness, yielding a dark brown powder. It was heated at 60℃ for 16 h, filtered through a DIAION HP-20 column, washed with water and eluted with 40% (v/v) ethanol. Evaporation of the eluate yielded a reddish brown powder containing the monomeric and oligomeric proanthocyanidin mixture, oligonol. The characteristics of the oligonol preparation used in this study (Batch No. OLF1202S) are shown in Table 1 [12]. Oligonol was dissolved in dimethyl sulfoxide (DMSO) and diluted to an appropriate concentration in culture media.
Cell culture
C6 glial cells (Korean Cell Line Bank, Seoul, Korea) were maintained in a culture flask containing 10% fetal bovine serum supplemented with Dulbecco's Modified Eagle's Medium (Welgene, Daegu, Korea) (pH 7.2) at 37℃ in a humidified atmosphere of 5% CO2 in air. All subsequent procedures were performed under these conditions. The cells were subcultured with 0.05% trypsin-EDTA in phosphate buffer saline.
Induction of oxidative stress
After confluence had been reached, the cells were seeded into 96-well plates at 5 × 104 cells/mL and allowed to adhere for 2 h. Next, cells were treated with sodium nitroprusside (SNP) (500 µM) or hydrogen peroxide (H2O2) (500 µM). After 24 h of incubation, samples were treated in the test wells at various concentrations for 24 h.
Cell viability
Cell viability was assessed using the MTT colorimetric assay. MTT solution (5 mg/mL) was added to each 96-well culture plate, followed by incubation for 4 h at 37℃; the medium containing MTT was then removed. The incorporated formazan crystals in the viable cells were solubilized with 200 µL of DMSO, and, after 30 min, the absorbance of each well was read at 540 nm using a microplate reader (Model 680, Bio-Rad, Hercules, CA, USA).
NO level
The amount of NO production was assayed by measuring the accumulation of nitrite, using a microplate assay method based on the Griess reaction. Briefly, 100 µL of culture supernatant was allowed to react with 100 µL of Griess reagent (Sigma-Aldrich Co. St. Louis, MO, USA), followed by incubation at room temperature for 15 min. The optical density of the samples was measured at 540 nm using a microplate reader.
Reactive oxygen species (ROS) generation
After 24 h of sample treatment, 2',7'-dichlorofluorescein diacetate (DCFDA) (Sigma-Aldrich Co. St. Louis, MO, USA) (80 µM) was added, and the cells were incubated for 30 min at 37℃. Then, fluorescence intensity was measured at 485 nm excitation and 535 nm emission using a fluorescence spectrophotometer (FLUOstar OPTIMA, BMG Labtech, Ortenberg, Germany).
RNA extraction and Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was isolated using Trizol reagent (Invitrogen Co., Carlsbad, CA, USA) by following the manufacturer's methods. The total RNA was digested with RNAse-free DNase (Roche, Indianapolis, IN, USA) for 15 min at 37℃ and re-purified using the RNase kit according to the manufacturer's protocol (Qiagen, La Jolla, CA, USA). cDNA was synthesized from 2 µg of total RNA by incubation at 37℃ for 1 h with AM reverse transcriptase (Amersham, Arlington, USA) and random hexanucleotides according to the manufacturer's instructions. The following primers were used to amplify the specific genes of interest: forward, 5'-TTC-AAA-TCA-GAT-TCT-GGG-AAA-AT-3' and reverse, 5'-AGA-TCA-TCT-CTG-CCT-GAG-TAT-CTT-3' for the COX-2 gene; forward, 5'-AGA-GAG-ATC-CGG-TTC-ACA-3' and reverse, 5'-CAC-AGA-GCT-GAG-GGT-ACA-3' for the iNOS gene; and forward, 5'-GCA-GCC-TAT-CAC-CAA-CTC-3' and reverse, 5'-TAC-TCC-TTC-TTC-TCC-ACC-3' for the NF-κB p65 gene.
RESULTS
Protective effect of oligonol against oxidative stress induced by SNP
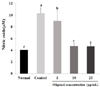
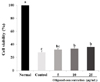
SNP was applied for induction of NO in C6 glial cells and the protective effects of oligonol were investigated. Fig. 1 shows that the control produced 10.34 µM of NO by SNP, two times more than the normal group. However treatment with oligonol at a concentration of 5 and 10 µg/mL led to a decrease in formation of NO to 9 µM and 4.5 µM, respectively. This result indicated that SNP induced overproduction of NO, whereas oligonol significantly inhibited the production of NO. As shown in Fig. 2, the cell viability of the control group, which was only treated with SNP, was 28.23%, indicating that cells died as a result of oxidative stress induced by SNP. Treatment with oligonol resulted in a significant increase of cell viability in a concentration dependent-manner.
Protective effect of oligonol against oxidative stress induced by H2O2
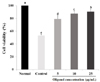
Fig. 3 shows the protective effects of oligonol on oxidative stress induced by H2O2. The cell viability of the control group showed a significant decrease to 43.2% compared to the normal group, with 100%. C6 glial cells were damaged by treatment with H2O2. However, treatment with 5 and 10 µg/mL concentrations of oligonol led to an increase of cell viability to 79.2% and 87.5%, respectively. In particular, at the concentration of 25 µg/mL, the cell viability was recovered to almost normal level.
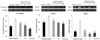
Fig. 4 shows the scavenging effect of ROS of oligonol determined by DCFDA assay. The control group showed a steady increase of ROS formation compared to the normal group. Based on these data we can predict that oxidative stress is induced by increases of ROS formation. However, treatment with oligonol at a concentration of 5, 10, and 25 µg/mL led to a decrease in ROS formation to 89.7%, 87.1%, and 86.4%, respectively. This result indicates that oligonol inhibited ROS formation in a concentration-dependent manner.
Protective mechanism of oligonol against oxidative stress
To investigate the protective mechanism against oxidative stress and inflammation, we observed the effect of oligonol on mRNA expression of NF-κB p65, COX-2, and iNOS in H2O2 treated C6 glial cells (Fig. 5). Control group treated with H2O2 showed higher expression of mRNA in NF-κB p65 compared with the normal group. However, treatment with 5 and 10 µg/mL oligonol led to decreased expression of NF-κB p65. At a concentration higher than 10 µg/mL, NF-κB p65 mRNA expression was down-regulated to almost the normal level. COX-2 mRNA expression showed a pattern form similar to that of NF-κB p65. Treatment of glial cells also increased the expression of COX-2 mRNA. However, oligonol had a downregulatory effect against overexpression of COX-2 mRNA. In addition, marked up-regulation of iNOS was observed by H2O2 treatment, whereas oligonol induced a significant decrease in iNOS expression. Combining all results together, oligonol regulated mRNA expression related to oxidative stress and inflammation such as NF-κB p65, COX-2, and iNOS.
DISCUSSION
Oligonol is a low molecular weight polyphenol in Litchi chinensis Sonn. Oligonol contains catechin-type monomers and oligomers derived by a novel manufacturing process of the Litchi chinensis Sonn. [14]. Several studies have reported on the antioxidative and anti-inflammatory potential of oligonol [13,15]. These results support the capacity of oligonol in regulating the pathological condition of chronic disease, such as cancer, diabetes, cardiovascular disease, and neurodegenerative disorders as well as aging progression. It has already been shown that oligonol is absorbed from the intestine, and has a strong antioxidative activity and anticancer effects. Based on these evidences, we could expect that oligonol might have protective effects against neurodegenerative diseases induced by oxidative stress. However, study on the protective effect of oligonol against oxidative stress in glial cells related to cognitive and memory impairment has not yet been conducted.
The number of people suffering from neurodegenerative disease is increasing. The search for functional food or bioactive compounds against neurodegenerative disease is crucial for prevention and treatment of neurodegenerative disease. The oxidative stress and inflammatory process induced by free radicals are the main cause of neurodegenerative disease. In particular, overproduction of NO causes damage to cells and is very important factor in neurodegenerative diseases. NO has several potential roles in the brain, as a neurotransmitter, a potential source of free radicals, and a potential mediator of inflammation [16,17]. Overproduction of NO can lead to react with membrane lipids to induce lipid peroxidation. Indirectly the combination of NO and O2- can form highly reactive intermediates, such as ONOO-, which can induce DNA strand breaks, lipid peroxidation, and protein nitration [18]. Injury of glial cells also occurred by NO through activation of microglia. Activated microglia produce a variety of pro-inflammatory and neurotoxic factors, including cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) [3,5,19,20]. In the current study, NO was induced by SNP in C6 glial cells and the effect of oligonol was investigated. C6 glial cells were used and oxidative stress was induced by SNP. As shown in Fig. 1, oligonol inhibited the production of NO, suggesting a protective role against NO-induced oxidative stress. As the concentration of oligonol increases, production of NO shows a progressive decrease, almost to the normal level. In addition, as shown in Fig. 2, treatment with oligonol resulted in elevation of cell viability compared with the control group. In this experiment we confirmed that SNP elevated NO production and inhibited cell growth. However, oligonol had a protective role against inflammation by inhibiting production of NO, resulting in elevation of cell viability.
H2O2 is one of the reactive oxygen species (ROS) involved in oxidative stress. By the high accumulation of oxygen radicals and H2O2, protein changes occur in intracellular and intercellular signaling pathways, so that proteins become inactivated or damaged, resulting in cellular degeneration and death [21]. The current results also support that treatment of C6 glial cells with H2O2 resulted in increased ROS production and decreased cell viability. However, oligonol inhibits ROS and it can protect brain cells from death induced by overproduction of ROS. By inhibiting inflammatory factors, oligonol can protect against glial cell death from oxidative stress and it would be related to improvement of memory and cognitive function. H2O2 regulates expression of inflammatory factor genes such as NF-κB, iNOS, and COX-2. NF-κB activation is essential for iNOS expression and NO regulates NF-κB at various points in its activation cascade. NO has a positive effect on NF-κB activation by affecting signaling cascades [22]. Activation of NF-κB culminates in translocation of NF-κB to the nucleus and an increase in gene expression such as iNOS and COX-2 [23,24,25]. iNOS has the ability to produce NO used during host defense. However, overproduction of NO is also injurious to host cells, leading to neurotoxicity and disease. iNOS gene expression is regulated through transcriptional controls [26]. Chemical stimuli that regulate NF-κB activity can also modulate iNOS induction [27,28]. COX-2 is activated by inflammation inducible factor such as IL-1, TNF, LPS, and reactive oxygen intermediates [29]. COX-2 induces inflammation by increasing the formation of eicosanoids. COX-2 increases the conversion of arachidonic acid into prostaglandin E2 (PGE2), which sets up a negative feedback loop because PGE2 acts through cyclic AMP to inhibit transcription [17]. Which inactivates proteins or cells leading to death. In this study, we suggested the theory that oligonol might have an effect on inflammatory factor gene which is regulated by H2O2. Oxidative stress was induced in C6 glial cells by H2O2; however, oligonol had a protective effect on inflammatory factor genes. Oligonol decreased expression of mRNA of NF-κB, iNOS, and COX-2. Several reports have also demonstrated the anti-inflammatory effect of oligonol [30,31]. Oligonol decreased pro-inflammatory cytokines (IL-1β, IL-6, TNF-α), COX-2 and iNOS in LPS-stimulated RAW 264.7 cells [30]. In addition, in the kidney of type 2 diabetic mice, inflammation-related genes and NF-κB transcription-stimulated AGE-RAGE interaction were down-regulated [31]. Together with this evidence, we suggest that oligonol regulates oxidative stress-induced inflammation.
The current results indicated that oligonol blocked the oxidative stress induced by H2O2 and also caused a decrease in expression mRNA of inflammatory factor, which has a connection with H2O2 mechanism. These results indicated that oligonol regulated the mechanisms related to oxidative stress and inflammation, eventually leading to elevation of cell viability.
Although further study on the protective mechanism of oligonol from oxidative stress is needed in order to support these findings, the current study suggests the promising role of oligonol as an antioxidant against oxidative stress in glial cells related to cognitive impairment.


 XML Download
XML Download