

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Adipocytes are known simply as energy reservoirs, but since the 1990s, studies have revealed that adipocytes directly and indirectly regulate body energy metabolisms by secreting a protein called adipocytokine [1,2]. Adipocytokines which includes leptin and adiponectin play a pivotal role in regulating energy metabolisms. Adipocytokine have been noted for causing metabolic disorders like obesity, diabetes, and arteriosclerosis, and thus, regulating the adipocytokine has been considered a novel approach in treating such metabolic disorders [3].
Differentiation of adipocytes is a complicated process affected by various hormones and transcription factors. Adipocyte differentiation is primarily regulated by various transcription factors such as peroxisome proliferator activated receptors (PPARs) and CCAAT/enhancer binding proteins (C/EBPs) [4]. These factors are expressed at different stages of differentiation process. PPAR expression is regulated by C/EBP family. Specifically, the C/EBP induces early expressions of the adipocyte differentiation of PPAR; and the PPARγ regulates the expressions of several adipocyte-secreted proteins including adiponectin [5].
Researchers have shown interests in developing natural ingredients that can prevent and manage obesity. Resveratrol is a phytochemical produced naturally by several plants when under attack by pathogens such as bacteria or fungi [6]. Resveratrol has been a topic of numerous animal and human studies into anti-aging effects or various chronic diseases preventions [7,8,9,10,11]. Resveratrol also triggers mechanism that counteracts obesity-related effects like restraining fat synthesis and accumulation, and resveratrol was shown to be more effective in high-fat diets than in normal diets [12,13].
Major food sources of resveratrols are skin of grapes, wines, and peanuts [6]. Studies have shown that the resveratrol content in peanut sprouts is 110.3 µg/g, which is much higher than grapes, 3 µg/g and wine, 0.6 µg/g and peanut, 0.018~1.25 µg/g [6]. Peanut sprouts refer to germinated peanut seeds. Peanut sprout is germinated and grown by spraying potable water under humid and warm condition and it can be harvested after 7 days from starting spray [12].
Peanuts itself contains resveratrol as little as 0.15 µg/mg while when peanut is germinated and grown into sprout, resveratrol is significantly increased to 14.2 µg/mg, as much as 90 times of peanuts [14]. Peanut sprout contains isoflavones such as daidzin, genistin, genistein and various types of polypenols [15]. Peanut sprout also includes essential fatty acids and essential amino acids, especially plenty of asparagine, which are far more than peanut itself [15].
Despite of these nutritional values of peanut sprout, the study determining physiological functions of peanut sprout is very limited, and most of them are in vitro studies regarding antioxidant ability of peanut sprouts [16,17,18]. In Korea, since bean sprouts is habitually consumed as a vegetable, there is a great possibility that peanut sprout can be used as a fresh vegetable as well as functional health foods Therefore, this study aims to find out the effects of peanut sprout extracts that contain large amounts of resveratrol on weight controls and protein expressions of transcription factors related to adipocyte differentiation and adipocytokine for rats under high-fat diets.
MATERIALS AND METHODS
Preparations of peanut sprout extract (PSE)
The peanut sprout extracts were prepared from Chonnam National University, by the modified methods of Wang et al. [19]. Briefly, fresh peanut sprout, which was germinated for 7 days with mature peanut (Arachis hypogaea L.) kernels, was obtained from a local processing plant (JangSuChae Co, Gyeonggi-do, South Korea). Peanut sprouts were cooked in a boiling water bath. After they were heated, peanut sprouts were cooled with tap water and subjected to lyophilization to prepare dried sprout powders. Dried peanut sprout was extracted with ethanol (1:10 v:v) at 60℃ for 90 min and filtered. The extraction procedure was repeated 3 times. Ethanol in the sample was removed using a rotary vacuum evaporator (N-11 Eyela, Tokyo Rikakikai Co. Tokyo, Japan). Resveratrol contents in PSE were determined by the modification of Wang et al's study [16]. The mean resveratrol content of PSE used in this study was 38.17 mg/mL.
Animals and Experimental diets
Four week-old Sparague-Dawley (SD) were assigned to two groups; 10 normal-fat (NF) diets, 7% fat diets, and 30 high-fat (HF) diets, 20% fat diets, according to a randomized block design. After 4 weeks, the high-fat diet group was divided into 3 groups; high-fat (HF), high-fat with low levels of peanut sprout extracts (HF + PSEL), and high-fat with high levels of peanut sprout extracts (HF + PSEH), and were all fed for another five weeks. Low peanut sprout extract diets (HF + PSEL) had 15 mg/kg (0.025% resveratrol) and high peanut sprout extract diets (HF + PSEH) had 30 mg/kg (0.05% resveratrol) (Table 1). Food and water were freely accessed to rats in a stainless steel cage, and the temperature of room was maintained at 24 ± 1℃. All experimental diets were stored at 4℃ in the refrigerator. Dietary intake was recorded twice a week and weighed once a week at a fixed time. All experiments were approved by the guidelines of Animal Testing Ethics Committee of Eulji University (EUIACUC 10-09).
Specimen preparation of various organs
After 9 weeks of feeding, the experimental animals were fasted for 12 hours and anesthetized with ethylether. Blood samples were drawn from hearts with a syringe after laparatomy. The blood sample was centrifuged for 30 minutes at 3,000 rpm, and the plasma was stored at 70℃ before use. Specimens taken from liver, kidney, spleen, epididymal fat, internal fat, kidney fat, and thymus were washed with saline solution, dried in a filter paper, and then weights were measured.
Lipid measurements in adipose tissue
Adipose tissues were collected from various sites such as epididymal fat, kidney fat, and abdominal fat. Total lipids in adipose tissues were extracted with chroloform:methanol (2:1, v/v) using a modified method described by Folch et al. [20]. After extraction, total lipids were determined as described by Frings and Dunn [21], with slight modifications. Total cholesterol and triacylglycerol were measured enzymatically using a commercial kit (Asan Pharmaceutical Co., Seoul, Korea).
Plasma AST (aspartate aminotransferase) and ALT (alanine aminotransferase)
Plamsa AST and ALT activity was determined by commercial assay kit (Asan Pharmaceutical Co., Seoul, Korea) and determined absorbance at 505 nm with spectrophotometer.
Western blot analysis for PPARγ, C/EBPα, adiponectin, and leptin
The protein expressions of PPARγ and C/EBPα, and adipocytokines including adiponectin and leptin were determined [13]. Adipose tissue previously stored at -70℃ was finely chopped, mixed with 1 mL of cold phosphate buffered saline (PBS), and centrifuged at 3,000 rpm for 10 minutes. The 400 µL of lysis buffer (Cell Signaling Technology, Boston MA, USA) was added and mixed for 30 minutes, and the mixture was centrifuged at 4℃ at 13,000 rpm for 15-30 minutes. Protein concentration was measured at 595nm by Bio-red method.
After protein was quantified, the sample was dissolved in SDS-PAGE, and transferred to immobilon™-p membrane (Millipore, Bedford, MA, USA). The membrane was 5% milk/TBST (20 mM Tris-HCl, 137 mM Nacl, 0.1% Tween 20, pH 7.4) and incubated at room temperature for 1 hour. Antibody of the protein was prepared by diluting PPARγ, C/EBPα, leptin (Santacruz, CA, USA), and adiponectin (Cell Signaling Technology, Boston, MA, USA) with 5% milk/TBST and then incubated. After being washed with TBST, the membrane was incubated with Peroxidase- Affinipure Sheep Anti-Mouse IgG (Jackson immunoresearch, West Grove, PA, USA) or Perox-Affinipure Donkey Anti-Rabbit IgG (Jackson immunoresearch, West Grove, PA, USA). Its color was developed by WEST-ZOL® Plus Western Blot Detection System (iNtRON Biotechnology, Korea) and X-Omat Film (Kodak, Tokyo, Japan) and the molecular weight was compared and analyzed with High-Range Rainbow™ Molecular Weight Marker (GE Healthcare, Uppsala, Sweden). Each band was measured for its density by an imaging program called image J Launcher (provided by NCBI).
Statistical analysis
The statistical analysis was performed using Statistical Analysis System (SAS Institute, Cary, NC, USA). The results were reported as mean with standard error, and statistically significant differences among groups were determined by using one way-ANOVA (analysis of variance). Statistically significant differences among the means of groups were tested at α = 0.05 using Duncan's multiple range tests.
RESULTS
The changes of weight and dietary intake
After 9 weeks of feeding, the NF group had an average weight gain of 311.1 ± 14.9 g in NF group, 349.7 ± 11.7 g in HF group, 352.1 ± 10.0 g in HF + PSEL group, and 333.5 ± 9.1 g in HF + PSEH group, showing HF + PSEH group have less weight gains than the HF group (Table 2). The total dietary intake after 9 weeks was 1,290.5 ± 42.8 g for NF group, 1,155.8 ± 23.3 g for HF group, 1,143.8 ± 18.0 g for HF + PSEL group, and 1,128.0 ± 24.6 g for HF + PSEH group. No significant differences among the HF group, HF + PSEL group, and the HF + PSEH group were shown.
The changes of organ weights
The liver weight of the HF group (22.0 ± 0.8 g), HF + PSEL group (22.6 ± 0.7 g), or HF + PSEH group (21.5 ± 0.8 g) were significantly heavier than the NF group (12.6 ± 0.6 g), but there was no significant difference between HF group, and HF + PSEH group (Table 3). Likewise, the kidney, and thymus weights of the HF group, HF + PSEL group, and HF + PSEH group were not significantly different.
The weight change of adipose tissues
Although HF + PSEH group showed the lower internal fat weight, 9.32 ± 0.8 g, than that the HF group, 10.97 ± 0.7 g, but there were no significant difference among groups (Table 4). HF + PSEH group had lowest epididymal fat weight, and the statistical difference between HF group and HF + PSEH group were shown (P < 0.05). The total fat weight between HF group and HF + PSEL or HF + PSEH was not statistically significant.
The lipid concentrations of adipose tissue
In Table 5, total lipids in the HF + PSEL group (228.1 ± 13.7 µmol/g) and HF + PSEH group (227.0 ± 9.8 µmol/g) decreased significantly compared to those in the HF group (240.5 ± 13.7 µmol/g) group (P < 0.05). Total cholesterol levels also decreased significantly in HF + PSEL group (6.6 ± 0.4 µmol/g or HF + PSEH group (6.5 ± 0.3 µmol/g) compared to those in the HF group (7.5 ± 0.3 µmol/g) (P < 0.05). No significant differences were observed for triglyceride levels among groups.
Plasma AST and ALT concentration
To determine the safety of peanut sprout extract as a supplementation, the plasma concentration of ALT and AST, as index of liver damage, were determined (Table 6). The plasma concentration of AST and ALT were increased when rats fed with high fat diet. PSEH supplementation caused AST and ALT levels to decrease, when it compared to HF group, but it was not statistically significant.
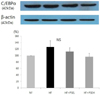
Effects of peanut sprout extracts on protein expressions of PPARγ and C/EBPα
By setting the NF at 100%, the effects of sprout peanut extracts on the protein expressions of PPARγ and C/EBPα in HF Groups were determined (Fig. 1, 2). The protein expression of PPARγ in HF + PSEH group was significantly lower than the HF group (P < 0.05, Fig.1). The protein expressions of C/EBPα in HF + PSEL group and HF + PSEH group were lower than that of the HF group, although the differences were not statistically significant (Fig. 2).
Effect of peanut sprout extracts on adiponectin and leptin
The protein expression level of adiponectin was significantly increased following the treatment with peanut sprout ethanol extracts at high levels (HF + PSEH group) when it compared with HF group (P < 0.05, Fig. 3). The HF group had 58.9%, HF + PSEL had 67.2%, and HF + PSEH had 76.2%. The protein expression of leptin in HF group was higher than that in NF group without statistical significances (Fig. 4). Although there was a much lower leptin level in the peanut sprout treated group than the HF group, the difference was not statistically significant.
DISCUSSION
World Health Organization (WHO) has announced obesity as a disease in need of treatment. Peanut sprouts refer to germinated peanut seeds and produce lots of resveratrol during germination [22]. In addition to abundant resveratrol contents in peanut sprouts, it also contains various bio-functional components such as pholypherols, isoflavones, and essential amino acids [15]. Hence, to evaluate peanut sprouts as a functional food material, anti-obesity effects of peanut sprout extract were examined by providing rats with high fat diet supplemented with low peanut sprout extract (0.025% resveratol) or high peanut sprout extract (0.05% resveratrol).
In this study, the body weight gain in high peanut sprout extract group was significantly suppressed, as compared with high fat group. However, the total food intakes were similar for the high fat groups and the peanut sprout extract groups. Likewise, Studies reported that hamsters provided with high fat diets added with resveratrol showed no significant differences in dietary intake and food efficiency ratio, but a significant decrease in weight [23,24].
In addition to decreased weight gains, there were also significant decreases in epididymal fat weights (Table 4) and low total lipids and cholesterol contents in adipose tissue of peanut sprout extract treated rats (Table 5), compared high fat only diet, which demonstrated clearly that peanut sprout has some anti-obesity effects. Many in vivo resveratol studies reported that 0.04% pure resveratrol administered into a high fat diet was effective in reducing body weight or decreasing adipose tissue weights while 0.01% resveratrol was not very effective [7,8,9,10,11,12].
In this study, the PSE supplementation (containing 0.05% resveratrol) showed significantly decreased visceral adipose fat weights, even much higher effects than pure 0.05 % resveratrol supplementation reported by previous studies [9,10,11,12]. Two previous in vitro studies reported that combination treatment with resveratrol and genistein or other natural polypherols suppressed lipid accumulation significantly more than the responses to resveratrol alone [25,26]. Those studies suggested that resveratrols's anti-adipogenic activity was substantially enhanced by the other natural isoflavornes or pholypherols compound in foods. Therefore, we believe that several biofunctional components of peanut sprout, in addition to resveratrol, allowed obtaining much stronger effects at reducing adipose body fat weights and body weight gain. This study has limitation that other physiological bio-components of peanut sprouts were not analyzed, and thus more studies regarding various components of peanut sprout on anti-obesity effects are needed.
In this study, the addition of the PSE supplementation in high fat diet lowered the epididymal fat weight even more so than the normal diet. A research found that the epididymal fat cell size of rats fed a normal diet with natural plants supplementation was significantly less than that of rats fed normal diet [27]. Based on our data of AST and ALT concentration, and the fact that adiponectin expression in adipose tissue significantly increased with the PSE supplementation, it can be concluded that the PSE supplement does not incur any side effects. However, further research may be needed to explore the causes for this decrease in epididymal fat weight.
Peanut sprout extract in this study also significantly decreased the expression of PPARγ (Fig. 1.), adipogenic transcription factor, and the target gene, adiponectin (Fig. 4.). These changes of PPARγ and adiponectin may contribute to the lower weight gain and epidermal fat weights in rats fed with high fat with peanut sprout extracts. In research on adipocyte differentiation in human and rats, the transcription factors such as PPARs and C/EBPs are reported to play an important role in adipocyte-specific gene expressions and adipocyte differentiations [4]. Vadal-Puig et al. [28] observed decrease in PPARγ expression due to restriated food intakes and increases in high-fat diets. Also, in a study of obese patients, it was found that PPARγ expression occurs mainly in human adipose tissues and is positively correlated with body mass index [29]. Our previous study which uses 3T3-L1 adipocytes [13] showed that resveratrol was down-regulated with adipocyte differentiation in a dose dependent manner.
Adiponectin is a direct target of regulation by PPARγ, and Tsuchida et al. reported that activation of PPAR increased adiponectin receptor 1 and 2 in both white adipose tissues (WAT) and brown adipose tissues (BAT) in vivo [30]. When treated with Pioglitazone, a PPARγ agonist, 3T3-L1 adipocytes presented enhanced insulin sensitivity through an upregulation of adiponectin receptor 2 (AdipoR2) and adiponectin [31,32]. In this study, high-fat diet groups (HF) showed significant decreases in protein expression of adiponectin, while peanut sprout extract diet groups showed significant increases in adiponectin. Adiponectin is a peptide hormone that is inversely related with abdominal visceral fat area, and diminishes the concentration of blood fatty acids and level of triglycerides [33,34]. Barnea et al. [35] concluded that adiponectin was decreased for mice fed with high fat diets and Rogers et al. [36] reported that rats fed with resveratrol showed increases in the concentrations of adiponectin in serum and suppressions of fat accumulation. Thus this can suggest that peanut sprout extract supplementation activates the protein expressions of PPARγ which inhibits adipocyte differentiation and improves adiponectin secretion in adipose tissues which has been shown to play important roles in lipolysis and thermogenesis.
Blood leptin concentration is also known to be closely related with body mass index and amount of body fat; an obese person tends to have higher blood leptin concentrations than a normal individual [37]. Szkudelska et al. [38] noted that resveratrol directly restrains leptin secretion by observing the effects of resveratrol in restraining leptin secretion in mice adipocytes. In this study, we observed the tendency of the peanut sprout extract groups to have lower leptin expression levels than the high-fat diet groups, though the difference was not significant.
In conclusion, the peanut sprout extract demonstrated decreasing weight gain and reducing body fat contents in adipose tissues of rats fed with high fat diets. Peanut sprout extracts can restrain adipocyte differentiation by lowering the expressions of PPARγ and adiponectin which are necessary for the adipocyte differentiation process.









 XML Download
XML Download