

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Inflammatory responses are recognized as the natural defence mechanisms that are critical for the recruitment of a variety of immune cells and molecules to the sites with the infectious microbes or the injured tissues [1]. Acute inflammation is a limited beneficial process, particularly in response to the infectious pathogens, whereas the chronic inflammation is an undesirable persistent phenomenon that can ultimately result in the development of inflammatory diseases [2,3]. Inflammation is an innate immune event that is primarily mediated by a variety of immune cells [4]. The inflammatory macrophages are generally induced by immunogens, which include various inflammatory molecules and inflammatory mediators that interact with the pattern-recognition receptors and their adaptor molecules [5]. Macrophages play a central role in producing the soluble factors, such as nitric oxide (NO), cyclooxygenase-2 (COX-2), and prostaglandin E2 (PGE2), as well as some cytokines in response to the extracellular stimuli including bacterial lipopolysaccharide (LPS) [6,7]. These factors are primarily controlled by the surface molecules, such as the pattern-recognition receptors (e.g., toll-like receptor 4) and their counter-adaptor molecules, which include TANK binding kinase 1, Toll-IL-1 receptor-domain-containing adapter-inducing interferon-β (TRIF), TRIF-related adaptor molecule (TRAM), and myeloid differentiation primary response gene 88 (MyD88) [8]. These inflammatory events require functional upregulation of the intracellular signaling machinery, including the levels of transcription factors and the upstream signaling cascades [9]. Thus, the suppression of the targets specific to the inflammatory process may have a great potential for preventing and treating the inflammation-mediated diseases.
Synurus deltoides (Aiton.) Nakai (Compositae) is wildly distributed in the mountainous areas and is one of the edible green-hued plants used as a natural coloring-agent for the rice cakes [10]. The plant has been used in the traditional medicine system to treat cystitis, bleeding, vomiting, hematemesis, and edema [11]. Several chemical compounds (e.g., anthocyanins, 20-hydroxyecdysone, terpenoids, coumarins, flavonoids, and triterpenoids) have been isolated from S. deltoides [12-15]. Despite the occurrence of some preliminary published works describing a variety of its pharmacological activities, including the antioxidant, antimutagenicity, and anti-inflammatory activities [16-18], the precise molecular mechanisms underlying the anti-inflammatory properties of the plant have not been fully investigated. In this study, we aimed to elucidate the anti-inflammatory activities of the ethanol extract of S. deltoides (SDE), using the LPS-activated macrophages and the acute in vivo inflammatory models.
Materials and Methods
Plant material and chemicals
The dried powder of S. deltoides leaves (10 kg) was extracted three-times with ethanol at room temperature for 24 h. Following the drying process, by the evaporation of water using a vacuum rotary evaporator, a crude extract of 974.58 g was produced. A voucher specimen (No. 2011SD) was deposited in the molecular plant biotechnology lab. The supplies of sulfanilamide, naphthylethylenediamine dihydrochloride, 2',7'-dichlorodihydrofluorescein diacetate (DCFH-DA), LPS (E.coli 0111 : B4), phorbal-12-myristate-13-acetate (PMA), and 1-(4,5-dimethylthiazol-2-yl)-3,5-diphenylformazan (MTT) were purchased from Sigma (St. Louis, MO, USA). The kit for RNA isolation and the first-strand cDNA synthesis were obtained from Invitrogen (Carlsbad, CA). The RPMI medium 1640, Dulbecco's modified Eagle's medium (DMEM), trypsin-EDTA, and fetal bovine serum (FBS) were acquired from Gibco BRL (Grand Island, NY, USA). All culture supplies were obtained from the BD-Falcon brand (BD, Franklin Lakes, NJ). The phospho-specific ERK, c-Jun N-terminal kinase (JNK), IκBα, p38 mitogen-activated protein kinase (p38 MAPK), and the total antibodies to ERK, JNK, IκBα, p38 MAPK, and β-actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The antibodies against NF-κB, c-Fos, and c-Jun were purchased from Cell Signaling (Beverly, MA, USA). Antibody-binding was detected with WEST-SAVE Up™ enhanced chemiluminescence (ECL) Western blotting substrate (AbFrontiers, Suwon, Korea). All other chemicals were of the analytical grade.
Cell line and cell culture
The RAW 264.7 cells and the human embryonic kidney cells (HEK 293) were purchased from the Korean Cell Line Bank (Seoul, Korea). The RAW 264.7 cells were maintained in RPMI 1640, supplemented with 10% FBS, 100 U/ml of penicillin, and 100 µg/ml of streptomycin. The HEK 293 cells were grown in DMEM, and supplemented with 10% FBS, 100 U/ml of penicillin, and 100 µg/ml of streptomycin. The cells were incubated at 37℃ in a humidified atmosphere of 95% air and 5% CO2.
Cell viability assay
The cytotoxicity of SDE on the RAW 264.7 cells was investigated. The cells were seeded into a 96-well plate at a density of 1 × 105 cells/well for 16 h and then exposed to the medium in the presence of different concentrations of SDE for 24 h. After removing the supernatant of each well, a total of 10 µl of the MTT solution [5 mg/ml in phosphate-buffered saline (PBS)] and 90 µl of FBS-free medium were added to each well at the time of incubation for 4 h at 37℃. The dark-blue formazan crystals formed inside the intact mitochondria were solubilized with 100 µl of MTT stop solution [containing 10% sodium dodecyl sulfate (SDS) and 0.01 M hydrochloric acid]. The amount of MTT formazan was qualified by measuring at 550 nm, using an enzyme-linked immunosorbent assay (ELISA) plate reader (ELx800TM, Bio-Tek, Winooski, VT, USA). The optical density of formazan formed in the control cells was taken as 100% viability. Cell viability was expressed as a percentage of the control culture value. Data were calculated as the percentage of inhibition by the following formula: Cell viability (%) = ODs/ODv × 100%. ODs and ODv indicated the optical density of cell lines incubated with SDE and vehicle control, respectively. Cytoprotective effect (%) = ODls/ODv × 100%. ODls and ODv indicated the optical density of cell lines incubated with SDE in present of LPS and vehicle control, respectively.
Determination of NO, PGE2, and TNF-α production
The RAW 264.7 cells were plated in a 96-well cell plate and stimulated with LPS (1 µg/ml) in the presence or absence of various concentration of SDE for 24 h. The aliquots of 100 µl of the cell culture medium were mixed with 50 µl of 1% sulfanilamide (in 5% phosphoric acid) and 50 µl of 0.1% naphthylethylenediamine dihydrochloride, at room temperature. The absorbance was determined at 550 nm using an ELISA plate reader (EL × 800TM). The levels of PGE2 and TNF-α were determined using the commercially available kits (Enzo Life Sciences, Farmingdale, NY), according to the manufacturer's instructions.
Intracellular ROS inhibition activity
A total of 5 × 104 RAW 264.7 cells were plated per well in a 96-well plate for 16 h. The cells were pretreated with various concentrations of SDE for 30 min, before being stimulated with LPS (1 µg/ml) for 24 h. After incubation, the supernatant of each well was removed, and the cells were washed with a preheated PBS at 37℃. DCFH-DA (20 µM) was then added, and the cells were incubated for 30 min. DCFH-DA was then removed from each well, and 100 µl of cold PBS was added. Fluorescence intensity (485 nm/535 nm, ex/em) was measured using a fluorescence spectrophotometer (Victor 3, PerkinElmer, New York, USA).
RNA preparation and reverse transcription-polymerase chain reaction (RT-PCR)
RT-PCR was used to analyze the gene expression in the RAW 264.7 cells, following stimulation with LPS in the presence of different concentrations of SDE for 6 h. The total RNA was isolated with a Trizol reagent in accordance with the manufacturer's instructions (Invitrogen, Carlsbad, CA). The first-strand cDNA was synthesized from the total RNA (2 µg), containing oligo (dT) primers and Moloney murine leukemia virus reverse transcriptase (M-MLV RT, Invitrogen). The primer sequences for iNOS, COX-2, TNF-α, and GAPDH are listed in Table 1. The aliquots of the individual PCR products were separated on 1% agarose gel, stained with ethidium bromide, and imaged using a Mini BIS image analysis system (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel). Densitometric analysis was done using an image analysis software (Quantity One; Bio-Rad, Hercules, CA, USA).
Luciferase reporter gene activity assay
The HEK 293 cells (1×106 cells/ml) were transfected with 1 µg of plasmids containing AP-1-Luc or NF-κB as well as β-galactosidase, using the PEI method in the 12-well plates. After 24 h, the transfected cells were treated with different concentrations of SDE in the presence or absence of PMA. The RAW 264.7 cells (1×106 cells/ml) were transfected with 1 µg of plasmids containing AP-1-Luc or NF-κB as well as β-galactosidase, using the lipofectamine 2000 in the 12-well plates. After 24 h, the transfected cells were treated with different concentrations of SDE in the presence or absence of LPS. Luciferase assays were performed using the luciferase assay system (Promega, Madison, WI, USA).
Cell lysis and imunoblotting
The RAW 264.7 cells (1 × 106 cells) were incubated in a 6-well plate. After 16 h incubation, the cells were treated with SDE for the predetermined times. Nuclear and total protein extracts were prepared [19]. The concentration of protein was determined using the Bradford assay. The aliquots of the lysates (40 µg of protein) were boiled at 94℃ for 5 min and separated on a sodium dodecyl sulfate-polyacrylamide gel and transferred to the polyvinylidene difuoride (PVDF; Bio-Rad) membranes. The membranes were blocked in a blocking buffer (Tris-buffered saline containing 3% BSA, 20 mM NaF, 2 mM EDTA, and 0.2% Tween 20) for 60 min at room temperature. The membrane was incubated for 60 min with the appropriate primary antibody at room temperature, washed three times with the TBST buffer (Tris-buffered saline containing 20 mM NaF, 2 mM EDTA, and 0.2% Tween 20), further incubated for 60 min with the HRP-conjugated secondary antibody, and washed three times with a TBST buffer. The bound antibodies were detected by the ECL system. Bands were finally visualized by a Mini BIS image analysis system (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel).
Animals
The ICR mice (6-8 weeks old, 17-21 g) were purchased from DAEHAN BIOLINK (Chungbuk, Korea) and housed at room temperature (24 ± 5℃), under relative humidity (50 ± 10%), with a 12 h light/12 h dark cycle. All animals were allowed free access to a commercial stock diet and water throughout the experiment. Studies were performed in accordance with the guidelines established by the Kangwon University Institutional Animal Care and Use Committee (KIACUC-12-0018).
Arachidonic acid-induced mouse ear edema
The ICR mice (n = 6) were orally pre-treated with SDE (40 mg/kg) or indomethacin (1 mg/kg) for 3 days. After the final treatment, arachidonic acid [2% (w/v)] was applied to the ear of the mouse (30 µl/ear). The ear thicknesses were measured using a dial thickness gauge (Mitutoyo, Japan), 1 h after the arachidonic acid treatment.
EtOH/HCl-induced gastritis
The inflammation of the stomach in mice was induced with EtOH/HCl according to a published method [20]. The fasted ICR mice (n = 6) were orally treated with SDE (100 mg/kg) or ranitidine (40 mg/kg), twice per day for 3 days. Thirty minutes after the final oral administration of SDE or ranitidine, the mice were orally treated with 0.3 ml of 60% ethanol in 150 mM HCl. Each animal was anesthetized with an overdose of ether, 1 h after the post-administration of EtOH/HCl. The stomach was excised, gently rinsed under the running tap water, inflated by an injection of saline, and then fixed for 30 min in 5% formalin. After opening the stomach along the greater curvature and spreading it out on a board, the area (mm2) of the mucosal erosive lesion was measured using a pixel-counter under a blinded condition. Ranitidine was used as a positive control drug.
Data analysis
All tests were carried out independently in triplicates (n = 3). The data are expressed as the mean ± standard deviation (SD). One-way analysis of variance (ANOVA) was used to determine the significant differences between the groups, followed by a Dunnett's t-test for multiple comparisons. A probability < 0.05 was considered as significant. All analyses were performed using SPSS 16 (SPSS Institute, Cary, NC, USA).
Results
Determination of cytotoxicity and NO, PGE2, and TNF-α production in RAW 264.7 cells
The cytotoxic effects of SDE were presented in Fig. 1A. The results showed that SDE alone at 100, 150, 200, 300, and 400 µg/ml did not exhibit any toxicity in the RAW 264.7 cells. SDL reduced cell viability to 68.54 ± 6.53% at 600 µg/ml (data not shown). Stimulation with LPS for 24 h led to a robust increase in the NO, PGE2 and TNF-α production. However, SDE significantly suppressed NO, PGE2, and TNF-α by the LPS-stimulated RAW 264.7 cells (Fig. 1B-D). Furthermore, this extract strongly prevented cells from the LPS-induced cytotoxicity at high concentrations (Fig. 1E).
Intracellular ROS inhibition activity
The levels of the ROS production in the RAW 264.7 cells were determined using a DCFH-DA fluorescence probe and a fluorescence spectrophotometer. As shown in Fig. 2, the ROS levels in the macrophages increased significantly to 532.40% when the RAW 264.7 cells were exposed to LPS, compared to the control without LPS. The pre-treatment with various concentrations of SDE rapidly suppressed ROS in the RAW 264.7 cells.
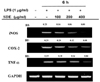
RT-PCR analysis of inflammatory-related genes
The iNOS, COX-2 and TNF-α mRNA expression in the unstimulated RAW 264.7 cells were undetectable; however, their mRNAs were profoundly induced after the treatment with LPS. Pretreatment with SDE markedly suppressed the LPS-stimulated iNOS, COX-2 and TNF-α expression (Fig. 3).
Luciferase reporter gene activity
As shown in Fig. 4, SDE significantly interrupted c-Jun, c-fos and p65 translocation to the nuclear fraction, without contaminating the cytosolic proteins, as confirmed by cytosolic β-tubulin (data not shown). Phorbol myristic acid (PMA) and LPS upregulated luciferase activity in the HEK 293 cells and RAW 264.7 cells, respectively, which were transfected with a luciferase reporter construct containing AP-1 or NF-κB binding sites. There were also suppressed by the SDE treatment (Fig. 5A-D).
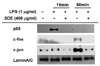
Imunoblotting analysis
As shown in Fig. 6A, at an early time point (5 min), the phosphorylation of p38 was suppressed by SDE. The ERK phosphorylation at 15 and 30min was also reduced by the SDE extract. However, SDE did not affect JNK phosphorylation. As shown in Fig. 6B, SDE blocked IκBα phosphorylation and Akt phosphorylation, an essential step for the NF-κB translocation. Furthermore, the phosphorylation of the upstream signaling kinase [p85 (a regulatory subunit of PI3K)] did not decrease following the SDE treatment (data not shown).
Arachidonic acid-induced mouse ear edema
As shown in Fig. 7, ear edema induced by the arachidonic acid treatment was significantly suppressed by SDE as well as in the positive control (indomethacin). However, the oral administration of indomethacin severely reduced the normal body weight, whereas SDE did not (data not shown).
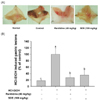
EtOH/HCl-induced gastritis
As shown in Fig. 8, the oral administration of EtOH/HCl dramatically increased the prevalence of the inflamed lesions in the stomach of mouse. The SDE (100 mg/kg) extract markedly reduced the gastric damage induced by EtOH/HCl to a level similar to the treatment with ranitidine (40 mg/kg).
Discussion
Inflammation is generally recognized as a cause of various diseases, such as cancer, diabetes, atherosclerosis, sepsis, and obesity. The inflammatory processes are mediated by multiple molecular mechanisms [21]. Thus, the RAW264.7 macrophages provide a useful model for evaluating the anti-inflammatory agents. A number of different inflammatory mediators including NO, PGE2 and TNF-α are generated by the macrophages upon stimulation with LPS (a primary component of the Gram-negative bacteria cell wall) and plays an important role in the immunepathology of the acute or chronic inflammatory diseases [22,23]. This observation could lead to the development of novel anti-inflammatory drugs without side-effects, aimed at preventing such diseases. First, the viability was tested at various SDE concentrations in the cells by the mitochondrial reduction of MTT, and these values were expressed as a percentage of the viability with respect to the control group (100% viability) (Fig. 1A). Based on these results, the SDE concentrations less than 400 µg/ml were selected for further studies. The cells were treated with different concentrations of SDE for 30 min, followed by a 24 h treatment with LPS, NO, and PGE2. The TNF-α levels in the cell supernatants were determined by the Griess assay and an ELISA kit.
Mammalian cells are constantly exposed to ROS as a result of normal metabolic processes occurring during aerobic respiration; however, excessively high levels of free radicals or ROS generate oxidative stress lead to enhanced lipid peroxidation and oxidative stress in cells. The macrophages stimulated with LPS generate ROS via activation of a membrane-bound NADPH oxidase [24,25]. Therefore, SDE acts as a potential ROS scavenger in an oxidative environment to balance the ROS levels and may inhibit the cytotoxicity induced by LPS.
It remains unclear whether the SDE-mediated inhibition of NO, PGE2, and TNF-α is the consequence of inhibiting iNOS, COX-2, and TNF-α at the transcriptional level or due to some other mechanisms. Our studies were extended to determine the iNOS, COX-2, and TNF-α mRNA expression levels. The results suggested that the suppressive activity of SDE on iNOS, COX-2 and TNF-α were mediated via transcriptional levels (Fig. 3).
The LPS-induced transcriptional control of inflammation from the activated macrophages is mainly managed by the redox-sensitive transcription factors, such as NF-κB and AP-1 [26]. We next investigated which signaling events and transcription factors were targeted by SDE using the immunoblot analyses and the reporter gene assays, to better understand the mechanisms underlying the inhibited LPS-induced cytokine production. These results suggested that the nuclear translocation pathway of NF-κB and AP-1 for the transcriptional activation of the inflammatory genes could be targeted by SDE.
The MAPK and PI3K/Akt signalling pathways play critical roles in regulating cellular proliferation, survival, and differentiation. The pathways also control the pro-inflammatory mediator synthesis and release by the activated macrophages during the inflammatory response and coordinate the induction of many genes encoding the inflammatory mediators [27,28]. Therefore, the signalling pathway is an attractive target in the anti-inflammatory drug research. The downstream signaling events that induce nuclear translocation of these transcription factors have been elucidated (Fig. 3-4). Therefore, MAPKs and PI3K/Akt pathways were selected to evaluate whether SDE was able to modulate the upstream signaling events. As the AP-1 translocation is mediated by MAPK phosphorylation, we next investigated the effects of SDE on the LPS-stimulated phosphorylation of ERK, JNK, and p38 MAPKs in the RAW 264.7 cells. Some studies have shown that p38, ERK or IκBα leads to an activation of the transcription factors (e.g., NF-κB and AP-1) and suggested that the MAPKs and PI3K/Akt signalling pathways directly affected the inflammatory protein expression [29,30]. These results suggest that SDE may block the LPS-induced expression of the pro-inflammatory cytokines by inhibiting the Akt and MAP kinase pathways.
The topical application of arachidonic acid on the mouse ears and the EtOH/HCl-induced acute gastritis models are well-known models for the induction of in vivo inflammatory symptoms [31,32]. We next explored whether SDE could ameliorate ear edema and acute gastritis in mice, induced by arachidonic acid and EtOH/HCl, respectively. The oral administration of SDE (40 mg/kg) or indomethacin (1 mg/kg) was conducted for 3 days; and arachidonic acid was applied to the ears of mice. The data suggest the SDE extract having displayed a very strong in vivo efficacy in the various inflammatory models and may be applied in the oral drugs targeting various inflammatory diseases.
In summary, we demonstratedthat SDE significantly inhibited cytokine production via down regulation of iNOS, COX-2, and TNF-α mRNA expression in the LPS-stimulated RAW 264.7 cells. Furthermore, SDE protected against the LPS-induced cell death induced by ROS via the anti-oxidative effects. In particular, the SDE extract suppressed multiple pathways of the inflammatory signaling cascades, such as Akt, p38, and ERK, which were associated with the inactivation of AP-1 and NF-κB. The in vivo tests showed that the orally administered SDE ameliorated the acute inflammatory symptoms in the EtOH/HCl-induced gastritis and arachidonic acid-induced ear edema in mice. Therefore, it is suggested that SDE may be further developed in the novel anti-inflammatory remedy research. Additional studies are currently underway to identify the specific phytochemicals responsible for the anti-inflammatory activities of SDE.






 XML Download
XML Download