

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Insulin-like growth factor (IGF)-1 plays an important role in the regulation of cell proliferation, survival, and growth. Its primary action is mediated by binding to the insulin-like growth factor-1 receptor (IGF-1R). Activated IGF-1R via IGF-1 phosphorylates and activates the insulin receptor substrate-1 (IRS-1) and leads to the phosphorylation of Akt [1]. Akt subsequently activates the mammalian target of rapamycin (mTOR) and S6 kinase (S6K), which is involved in protein synthesis and cell growth. Autophosphorylated IGF-1R also activates Ras, Raf, MEK and ERK signaling pathways. The elevated level of plasma IGF-1 is associated with the risk of several cancers in multiple organs, such as the colon, prostate, and breast [2-4]. Moreover, the elevated tissue levels of IGF-1 enhance susceptibility to chemically induced skin tumorigenesis, and inhibition of IGF-1 signaling decreases tumor formation [5].
Many epidemiological and experimental studies have reported that natural dietary compounds found in fruits and vegetables reduce cancer incidences, as well as tumor multiplicity [6-8]. Quercetin is one of the most common flavonoids, and has potent anti-carcinogenic effects derived from its anti-inflammatory and anti-oxidative activities [6]. The amount of quercetin intake was negatively correlated with the incidence of colorectal [7] and lung cancers in clinical studies [8]. It has also been reported that quercetin inhibits hydrogen peroxide-induced DNA damage and enhances DNA repair in the colon cancer cell line [9]. Quercetin also induces apoptosis by activating the pro-apoptotic proteins, such as caspase 3 and caspase 9, and inducing cell cycle arrest in several cancer cells [10]. Most studies thus far, however, reported anticancer activities of quercetin in the context of its antioxidative properties, and only a limited number of studies have focused on its modulation of signaling pathways, such as NFκB, MAPK, p53, androgen receptor and estrogen receptor in tumor growth [11-14]. In this study, quercetin effectively inhibited the development of skin tumor in BK5. IGF-1 transgenic (Tg) mice and decreased the phosphorylation of IGF-1R, IRS-1, Akt and S6K in skin cancer cells, which indicates that quercetin has potent anticarcinogenic activity through the inhibition of IGF-1 signaling pathway. To the best of our knowledge, this is the first study to demonstrate that quercetin specifically blocks IGF-1R activation in vivo.
Materials and Methods
Materials
7,12-Dimethylbenz(a)anthracene (DMBA) and 5-bromo-2'-deoxyuridine (BrdU) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Tetradecanoyl phorbol-13-acetate (TPA) was purchased from Alexis Biochemicals (San Diego, CA, USA). All other chemicals were obtained from Sigma-Aldrich, unless otherwise indicated.
Animals and diet
BK5.IGF-1 Tg mouse were imported from Dr. John DiGiovanni's Lab (University of Texas M.D. Anderson Cancer Center, TX, USA) and bred in Catholic University of Daegu. All animals were kept in controlled conditions of humidity (50 ± 10%), light (12-hour light/dark cycle), and temperature (23 ± 2℃). Animal protocols utilized in this study were approved by the Animal Care and Use Committee at Catholic University of Daegu (IACUC-2010-8). The composition of AIN-93G diet (Control) or quercetin diet (AIN-93G diet containing 0.02% quercetin) was indicated in Table 1. Dose of quercetin was chosen based on a previous animal study, which has shown a decreased intestinal polyp multiplicity with 0.02% quercetin diet [15]. Food intakes were recorded daily and body weights were recorded once a week.
Animal Experiment 1 (Two-stage mouse skin carcinogenesis)
The first animal study was performed to investigate the consequences of quercetin diet on skin carcinogenesis. Seventeen male BK5.IGF-1 Tg mice were randomly divided into two groups; control (n = 8) and quercetin (n = 9) group. At 8 weeks of age, dorsal skins of the mice were shaved two days before tumor initiation. Initiation was accomplished by a single topical application with 200 nmol (50 µg) DMBA under a subdued light. At 2 weeks after initiation, the mice were treated with 6.5 nmol (4 µg) of TPA in 200 µl of acetone twice a week for 20 weeks. Tg mice were fed AIN-93G diet (control) or quercetin diet (AIN-93G diet containing 0.02% quercetin) at 10 weeks of age with TPA promotion until the end of the experiment. Tumor incidence (number of mice bearing tumors/total number of mice) and tumor multiplicity (number of papillomas/mouse) were manually counted and recorded weekly. Tumor size was measured in diameters and length of each tumor by a caliper once a week.
Animal Experiment 2 (Histological analysis of Epidermal cell proliferation)
Eight-week old twelve male BK5.IGF-1 Tg mice were divided into the control (n = 6) and quercetin (n = 6) groups to investigate the effects of quercetin diet on epidermal cell proliferation of the Tg mice. Animals were fed ad libitum with either a control or quercetin diet for 4 weeks. In each diet group, dorsal skins of the mice were shaved and treated with either acetone (n = 3) or TPA (6.5 nmol (4 µg)/200 µl acetone/mouse, n = 3) twice a week for 2 weeks before sacrificing the animals. For measuring the rate of epidermal cell proliferation, 100 mg/kg body weight of BrdU was injected i.p. into mice 1 hr before sacrifice. Dorsal skin was isolated from the mice, fixed in formalin, and processed for paraffin embedding. Tissue sections were stained with hematoxylin and eosin (H&E) solution and immunostained with antibodies against BrdU (BD Biosciences, San Jose, CA, USA). The number of BrdU-positive cells in the interfollicular epidermis was counted in five random tissue sections in each mouse. The labeling index was calculated as the percentage of BrdU-positive basal cells to the total number of basal cells in the interfollicular epidermis.
Cell Culture
The murine papilloma cell line, the MT1/2 cells [16], were kindly provided by Dr. Susan M. Fischer's Lab (University of Texas M.D. Anderson Cancer Center, TX, USA), and were maintained in Minimum essential medium (GIBCO, Carlsbad, CA, USA) containing 10% FBS and 1% penicillin-streptomycin. All cell lines used in this study were maintained in a 37℃ humidified incubator with 5% CO2.
MTT Assay
To assess cell viability and cell proliferation, the MT1/2 cells (1.25 × 104 cells/well) were plated in 96-well plates. The cells were treated with various concentrations of quercetin (10, 20, 30, 40 and 50 µg/ml) or IGF-1 (20 ng/ml) for an indicated time. Then, the medium was aspirated from the wells, and 100 µl fresh serum-free medium containing MTT reagent (1 mg/ml) was added to each well. The cells were incubated for 4hr at 37℃ and lysed by addition of 100 µl DMSO, and the optical density (OD) at 540 nm was then measured.
Western Blotting Analysis
Cells were harvested in a mild-lysis buffer (10 mM Tris-HCl (pH7.5), 100 mM NaCl, 1% NP-40, 50 mM NaF, 2 mM EDTA (pH 8.0), 1 mM DTT, 1 mM PMSF, 10 µg/ml leupeptin and 10 µg/ml aprotonin). Cell lysates were subjected to 10% SDS-polyacrylamide gel electrophoresis, and subsequently, transferred onto the polyvinylidene difluoride membrane. After transfer, the membrane was blocked with a 5% non-fat dried milk in 0.1% Tween-20-Tris-buffered saline (TTBS) and probed with antibody against phospho Akt (S473), Akt, phospho S6K (T389), S6K, phospho Erk1/2 (Thr202/Tyr204), Erk, pIGF-1R (T1131), IGF-1R, pIRS-1 (S307), IRS-1, pPTEN (S380/T382/383), and PTEN (Cell Signaling Technology, Danvers, CO, USA) at 1:1000 dilution for O/N at 4℃. After three times of washing with a TTBS buffer, the membrane was incubated with a horse radish peroxidase-linked secondary antibody (1:1000) and visualized by ECL kit (Bio Science Technology, Pohang, Korea) according to the recommended procedure.
Results
Dietary quercetin supplementation delayed the time of tumor incidence and suppressed tumor multiplicity in BK5.IGF-1 Tg mice
The effect of quercetin on skin carcinogenesis was investigated in vivo using BK5.IGF-1 Tg mice in the first animal experiment (experiment 1). The mice were grouped into the control and quercetin diet groups, and fed with each diet for 20 weeks with TPA promotion. During the experimental period, food intakes of the control and quercetin groups were 7.3 and 7.4 g/day, respectively, and there was no significant difference between the two groups (Table 2). The body weight was gradually elevated without significant differences between the control and quercetin groups (Fig. 1). In DMBA-TPA two stage skin carcinogenesis protocol, animals in the control group developed skin papillomas at 2 weeks and reached up to 100% tumor incidence at 4 weeks of TPA promotion (Fig. 2A). In the quercetin group, skin papillomas were developed at 4 weeks and 100% of tumor incidence was recorded at 6 weeks of TPA promotion. This data indicates that the incidence of tumors was delayed by 2 weeks by quercetin supplementation. Quercetin also decreased mouse skin tumor multiplicity by 35% compared to that of the control group (Fig. 2B). It has been reported that BK5.IGF-1 Tg mice spontaneously developed skin papillomas due to highly activated IGF-1 signaling pathways [5]. We observed that spontaneous tumor development in the DMBA-TPA untreated region of the skin was also reduced by quercetin supplementation (Fig. 2C, 2D).
Quercetin decreased total number and size of tumors
In animal experiment 1, we further investigated how quercetin changed the number and size of the tumors. Quercetin supplementation showed a significant decrease in the tumor diameter (length and width) compared to that of the control group. At the end of the experimental period, the average number of tumors < 0.5 cm was 97 and 67.2, 0.5-1 cm were 32.7 and 18, and > 1 cm were 2.8 and 0.8, in the control and quercetin groups, respectively (Table 3). Quercetin supplementation also decreased the total number of tumors in the quertcetin group compared to that of the control group (Table 3).
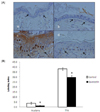
Quercetin reduced TPA-induced epidermal cell proliferation in BK5.IGF-1 Tg mice
To see the effect of quercetin on epidermal cell proliferation, we conducted a BrdU incorporation experiment (Experiment 2). TPA treatment induced skin hyperplasia in the control group, but quercetin supplementation remarkably reduced TPA-induced skin hyperplasia (Fig. 3A). BrdU labeling index was also reduced by quercetin supplementation, both in acetone and TPA treated animals (Fig. 3B).
Quercetin suppressed IGF-1 signaling by inhibition of IGF-1R phosphorylation in skin cancer cells
To investigate a mechanism on the inhibition of skin tumor development in BK5.IGF-1 Tg mice by quercetin, we analyzed the phosphorylation of protein components in the IGF-1 signaling pathway by a quercetin treatment in a murine papilloma cell line, the MT1/2 cells. As shown in Fig. 4A, IGF-1 stimulation significantly increased the phosphorylation of IGF-1R, IRS-1, Akt, and S6K, and 1 hr treatment of quercetin effectively suppressed the phosphorylation of these proteins in a dose dependent manner without affecting the cell viability (Fig. 4B). Phosphorylation of PTEN, a negative regulator of IGF-1 signaling [19], however, was not changed by the quercetin treatment. We further investigated whether quercetin could abrogate the IGF-1 induced cell proliferation. Simultaneous incubation of quercetin (10-50 µM) and IGF-1 for 48 hrs significantly inhibited the IGF-1 stimulated cell proliferation, and especially > 30 µM of the quercetin inhibited cell proliferation as low as the cells in a serum free media (Fig. 4C).
Discussion
Dysregulation of the IGF-1/IGF-1R system has been implicated in the proliferation of numerous cancer cells, including the colon, gastric, esophageal, hepatocytes and pancreatic cells, and a blockade of IGF-1 signaling pathways effectively inhibits tumor progression [2,3,17]. Commercially available compounds, inhibiting the IGF-1 signaling pathway (e.g.,wortmannin, picropodophyllin, NVP-AEW541), have been extensively investigated as a therapeutic option for people with malignant cancer [18,19]; however, it would be beneficial to offer natural therapies that could be easily incorporated into the diet and would act on the same target to reduce the risk of cancer. Particularly, quercetin highly abundant in plant food, such as onion, kale and broccoli [20,21], has been shown to have potent anti-carcinogenic and anti-tumorigenic properties in many organs, and suppress the growth and metastasis of the melanoma cells [12,13,22]. Although many studies reported the effects of quercetin in regulation of key elements in the cellular signal transduction pathways linked to the cell survival and proliferation, its molecular action in direct modulation of the IGF-1 signaling pathway remains to be answered.
The present study investigated a mechanism underlying the anticarcinogenic effect of quercetin on skin cancer in the context of IGF-1 signaling. BK5.IGF-1 Tg mice, which overexpress IGF-1 in the basal cells of skin epidermis, were utilized to confirm the effect of quercetin on the suppression of the IGF-1 signaling pathway. Quercetin supplementation decreased the DMBA-TPA induced tumor formation, as well as tumor size, and decreased TPA induced skin hyperplasia in Tg mice. Similar findings with epigallocatechin gallate (EGCG) have also been reported to decrease TPA induced skin tumor formation [23] and tumor size in lung cancer animal models [24]. BK5.IGF-1 transgenic mice also spontaneously induce tumors without TPA treatment due to spontaneous stimulation of IGF-1 signaling, and quercetin supplementation even suppressed spontaneous tumor formation. Although this study did not show the level of circulating IGF-1 in transgenic mice, IGF-1 has known for activating IGF-1 signaling pathway and inducing cell proliferation by autocrine or in a paracrine manner [25]. These data indicate the direct modulation of the IGF-1 signaling pathway by quercetin.
In addition to using an animal study to confirm the suppression of IGF-1 signaling induced skin tumor by quercetin, we analyzed the components of the IGF-1 signaling pathway in skin cancer cells to identify an actual target responsible for the chemopreventive effects of quercetin and elucidate a molecular mechanism of action. Quercetin treatment dose dependently decreased IGF-1 stimulated IGF-1R activation, as well as phosphorylation of IRS-1, Akt and S6K in the MT1/2 cells in this study. However, protein expression and phosphorylation of PTEN was not affected by a quercetin treatment. IGF-1R is a glycoprotein complex consisting of two extracellular α subunits, which confer binding specificity, and two transmembrane β subunits, which contain tyrosine kinase activity [26] and mediates action of IGF-1 on cellular differentiation and proliferation in cancer cells [27]. A ratio of IGF-1R and IRS-1 is mainly associated with cellular response to IGF-1 due to the inhibitory action of IRS-1 on IGF-1R induced cell differentiation [28]. The tumor suppressor gene, PTEN, is a negative regulator of the PI3K-Akt cell survival pathway, and activation of PI3K-Akt signaling through a loss of PTEN function is common in many other types of tumors [29-31]. However, protein expression and phosphorylation of PTEN was not affected by quercetin treatment, suggesting that suppression of Akt phosphorylation by quercetin is a PI3K cascade response of the suppression of upstream molecule of IGF-1 signaling. Based on the results of this study, IGF-1R appears to be the main target of quercetin in the blocking of IGF-1 signaling. It will be needed to further investigate whether quercetin directly binds to IGF-1R and inhibits its kinase activity.
On the other hand, how much of quercetin is absorbed and utilized at the physiological level is not consistent in the previously reported studies, since the bioavailability and metabolic range of quercetin (3-50 µg/L)[32] are varied by the type of supplements and food sources [33-35]. We used the doses of quercetin in the range of 10-50 µM in cell culture of this study and expect that these amounts can be achieved by eating plant based foods as a concentrated form, such as tea, tablet, and fruit skin extracts.
Signal transduction pathways in the downstream of IGF-1R are complex and have been studied extensively in cultured cells. Moreover, quercetin has been shown to inhibit a variety of enzymes involved in the cell proliferation and signal transduction pathways, including protein kinase C [36], tyrosine kinase [37], PI3K [38] and JNK [39]. Quercetin also has a binding activity to the type II estrogen binding sites [40] or aryl hydrocarbon receptors [41]. Therefore, other possible mechanisms of anticancer activity of quercetin cannot be excluded.
In this study, we identified that quercetin directly inhibited IGF-1R phosphorylation in the MT1/2 cells and suppressed skin cancer development in BK5. IGF-1 Tg mice. These results suggest a possible mechanism on how quercetin prevented cancer progression in the previous clinical and sub-clinical animal studies.






 XML Download
XML Download