

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Metabolic syndrome (MetS) increases cardiovascular morbidity and mortality and all-cause mortality [1]. The MetS is characterized by a constellation of interrelated metabolic risk factors that appear to induce the development of cardiovascular disease (CVD) [2]. The diagnosis of MetS is made based on the following clinical signs: central obesity, hyperglycemia, hypertension, and atherogenic dyslipidemia [3]. Atherogenic dyslipidemia consists of an aggregation of lipoprotein abnormalities including elevated triglyceride (TG) and apolipoprotein B and a reduced level of HDL cholesterol (HDL-C). Individuals with these characteristics commonly manifest a proinflammatory state [4]. Past prospective studies have shown an association between TG levels and coronary artery disease [5], and this association remained statistically significant after controlling for HDL-C and other risk factors [6]. More recent epidemiologic studies have reported that plasma TG and LDL particle size predicted subsequent coronary artery disease in 3 different Caucasian populations [7-9].
The primary goal of clinical management in individuals with MetS is to reduce the risk factors for clinical atherosclerotic disease. Because atherogenic dyslipidemia consists of abnormal levels of TG, small LDL particles, and reduced HDL-C, this condition can become a target for lipid-lowering therapy [3].
Recently, many vegetables, fruits, and herbs have been identified as protective agents against commonly occurring diseases. Epidemiological evidence suggests that high consumption of fruit and vegetables is associated with a reduced risk of cancer CVD [10,11]. These food products contain antioxidants, phenols, and reactive groups with protective properties [12]. Much of the supporting evidence for a preventive effect of polyphenols in various diseases is based on the finding that they facilitate both the scavenging of free radicals with concomitant formation of fairly stable radicals, and the chelation of transition metals, including iron [13,14]. A large number of studies suggest that these compounds have immunomodulatory and anti-inflammatory effects [15]. Therefore, there is a strong demand for natural bioactives obtained from plants.
Previous studies have shown that the bioactivity of functional food ingredients was significantly affected during processing and storage [16]. Kumamoto et al. [17] reported that Copper (II) ion increased the antioxidative activity of catechins in tea extract, but iron (II) ion largely inhibited them. Chen et al. [18] found that green tea contained 8 to 15 g of catechins in 100 g of dry leaves, whereas canned tea drink contained only 0.3-35 mg of total catechins per 100 mL. They suggested that the thermal degradation of catechins in various teas during processing, and loss of catechins during storage may occur. Factors that affect the stability of phenolic compounds include structure and concentration, pH, temperature, light, presence of co-ingredients, self-association, sugar and their degradation products, proteins, and sulfur dioxide [19,20]. In addition, the delivery of bioactive ingredients is challenging because of their limited solubility in water and off-odor.
Microencapsulation has been used for the preservation of natural components by entrapping the active ingredient in coating material [21,22]. Encapsulation involves formulating an active ingredient with secondary materials and preparation of a capsule that isolates the active ingredient from the environment until its release upon exposure to a desired stimulus [23]. Ersus and Yrudagel reported that microencapsulation using maltodextrin as the coating material gave the highest anthocyanin content at the end of the drying process [22]. Augustin et al. [24] reported that the microencapsulation of a bioactive mixture of fish oil, tributyrin, and resveratrol increased its bioavailability in the gastrointestinal tract and tissue distribution.
Although consumers are increasingly aware of the diet-related health benefits, a large group of the population lacks an adequate intake of fruits and vegetables. Thus, a dietary supplement such as a nutraceutical may be an alternative route to the consumption of functional ingredients with health benefits. This is particularly valid for phenolic compounds, which are not chemically synthesized and need to be extracted.
Rats fed high-fructose diets exhibit metabolic changes similar to those observed in MetS [25]. In the present study, we used a 65% high fructose diet to induce hypertriglyceridemia in rats, followed by an oral administration of green tea extract (GTE) or microencapsulated green tea extract (MGTE). In addition, we compared the effects of GTE and MGTE on lipid metabolism and cardiovascular tissue in hypertriglyceridemic rats.
Materials and Methods
Preparation of GTE and MGTE
GTE was obtained from Bioland Ltd. (Cheonan, Chungnam, Korea). The GTE contained 98% catechins, comprising 6 major ingredients including epicatechin gallated catechins, epicatechins, epigallocatechins gallates, gallocatechins gallates, and epicatechin gallates.
MGTE was prepared as follows: GTE was added into deionized water and maltodextrin (1:4, v/v) and then force-dispersed by hydroshear homogenizer (Janke & Kunkel, IKA® Labortechinik, Staufen, Germany) at 8,000 rpm for 30 min. Thereafter, the MGTE was dried by using a freezing dryer (SFD6M24L, Samwon Freezing Engineering Co., Seongnam, Korea).
Experimental animals and diets
A total of 40 male Wistar rats, 8 weeks of age, with an initial body weight of 247.2 ± 1.8 g (Orient-Bio laboratory Animal Research Center Co., Ltd., Seongnam, Korea) at the start of the experiment, were housed in individual stainless steel wire cages, in an animal room under constant conditions (12 h light-dark cycle, a temperature of 22 ± 1℃, and a relative humidity of 50 ± 5%). They had free access to food and drinking water. After a week of acclimation, the rats were randomly divided into 2 groups and fed either the AIN-93 diet (normal control group, NCON, n = 10) or the modified AIN-93 diet in which cornstarch was substituted with 65% fructose to induce hypertriglyceridemia, for 8 weeks [26]. Hypertriglyceridemia was defined as serum TG level > 200 mg/dL [27] by a LipidoCare Biosensor (MedicalSK, Inc., Daegu, Korea). Subsequently, the hypertriglyceridemic rats were randomly divided into 3 subgroups (n = 10 each) according to the serum TG levels. The 2 groups of rats were fed every day for 4 weeks either with GTE (hypertriglyceridemic GTE, HGTE) or with MGTE (hypertriglyceridemic microencapsulated GTE, HMGT) dissolved in distilled water using an oral Zonde needle (Natsume, Japan). In previous studies, the intake levels of GTE ranged from 50 to 120 mg kg·bw-1·d-1 [28,29]. In the present study, the daily dose of GTE or MGTE was set at 50 mg·kg·bw-1·d -1. The rats in the normal control group and in 1 high fructose fed group (hypertriglycridemic control, HCON) were fed with equal volume of distilled water. The ingredients used in this study were as follows: casein (Daejung Chemical and Metal Co., Gyenoggi-do, Korea); cellulose, mineral mix, and vitamin mix (G-Bio Co., Gyenoggi-do, Korea); cornstarch (Daesang Co., Seoul, Korea); sucrose (Samyang Co., Ulsan, Korea); fructose (Tate and Lyle, Decatur, IL, US); soybean oil (Cheiljedang Co., Seoul, Korea); and choline bitartrate and tert-butylhydroquinone (Sigma Aldrich, St. Louis, MO, USA). The animal experiments were conducted according to the accepted ethical rules for animal research and were approved by the Animal Ethics Committee of Kookmin University (Permission No. 201202).
Postprandial blood sampling and TG measurement
At the beginning and after the 8-week period on high fructose or normal control diet, food was removed from the rats at 09:00 am. At 14:00 pm, a fasting blood sample (20 µL) was obtained by tail-tipping and blood TG was measured using a LipidoCare Biosensor (MedicalSK, Inc., Daegu, Korea).
Blood and tissue sample processing
After 12 weeks of the experimental period, the rats were sacrificed following 16 h of starvation, by decapitation. The liver, heart, epididymal, and retroperitoneal fat pads were removed quickly, weighed, and stored at -70℃ until biochemical analysis. Blood samples were collected and centrifuged at 3,000 rpm at 4℃ for 15 min, and the supernatants were stored at -70℃ for biochemical analysis.
Biochemical analysis
The levels of TG, total cholesterol (TC), and HDL-C in the serum were measured by the enzymatic colorimetric method using commercial ASAN Reagents assay kits (Asanpharm Co., Gyenoggi-do, Korea).
The serum concentration of non-esterified free fatty acids (NEFA) and uric acid (UA) were analyzed using a commercial assay kit (EFFA-100; Bio-Assays Systems, CA, USA, UA; ASAN UA-Lq Reagent, Asanpharm Co., Korea) according to the manufacturer's instructions.
Enzyme activity analysis
The liver and retroperitoneal fat pads were homogenized in 0.25 M sucrose/Tris-HCl (0.01 M, pH 8) with a homogenizer (OMNI-INC, Marietta, USA), as described previously [30]. The homogenates were centrifuged at 40,000 × g for 10 min, and the supernatants were used to analyze enzyme activity. The assay medium for malic enzyme (ME) consisted of 100 mM Tris buffer (pH 7.4), 100 µL of 100 mM sodium malate, 50 µL of 20 mM, NADP+, and 0.75 mL of 20 mM MnCl2. The final volume of the assay mixtures was 3 mL in all cases. Enzyme activity was evaluated according to the increase in NADPH produced by the reaction catalyzed by malic enzyme [30]. The protein content of tissues was quantified by the SMART BCA Protein Assay Kit (21071; Intronbio, Korea)
Catalase activity was analyzed spectrophotometrically as described by Beers and Sizer [31]. The liver and kidney (0.4 g each) were homogenized with 1.6 mL of 50 mM phosphate buffer (pH 7.0). The homogenates were centrifuged at 10,000 rpm for 30 min, and then 2 mL of the supernatant was allowed to react with 1 mL of 30% hydrogen peroxide (H2O2). The decrease of 30% H2O2 in the presence of tissue homogenate was measured at 240 nm after 2 min.
Histomorphometric analysis
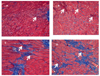
Heart tissue was fixed in 10% phosphate-buffered formalin for 1 day and processed in a routine manner to generate 5-µm-thick paraffin sections. Sections were stained with Masson-Trichrome staining and examined microscopically. Approximately 10 randomly selected fields of each heart sections were observed with a light microscope at 100 × magnification. The occurrence of myocardial fibrosis was examined in each heart section and the percentage estimated for every field. Myocardium thickness and lumen diameter of the left ventricle (LV) were measured with a ruler, and images were captured at 400 × magnification on a flat computer monitor. Five randomly selected images of the LV from each heart were used for measuring myocardial thickness and the diameter of LV was measured at the largest and the smallest part.
Statistical analysis
All data were analyzed using the statistical package for the social science (SPSS, version 20). One-way analysis of variance (ANOVA) was performed, and Duncan's multiple range test was used to determine the significance of differences among the groups. A P value of < 0.05 was considered statistically significant.
Results
Body weight, food intake and organ weight
The body weight, daily food intake, and organ weights are presented in Table 1. High fructose diet did not affect food intake. While feeding high fructose diet increased final body weight (407.7 g vs 429.8 g), GTE treatment suppressed the increase of final body weight over 4 week study. Although the significant difference between final body weights of HGTE and HMGT groups was not observed, the effect of GTE on body weight gain tended to be lower compared to that of MGTE treated rats. The administration of GTE reduced food intake in HGTE group significantly (P < 0.05), but the MGTE did not.
Significant liver weight increases were observed in high fructose fed rats with the mean increase in absolute liver weight of high fructose fed rats compared with controls being 40%. The administration of GTE and MGTE also produced statistically significant decreases in the weights of epididymal fat and retroperitoneal fat pad in high fructose fed rats.
Blood profiles
The initial mean postprandial serum TG concentration in the high fructose control diet-fed rats before GTE or MGTE treatment was 219.8 ± 77.5 mg/dL. Fig. 1 illustrates the changes in postprandial serum TG levels over a 4-week period of high fructose diet and high fructose diet with GTE or MGTE treatment. Fructose-fed rats had a higher serum TG level (P < 0.05) than normal control diet-fed rats and it remained elevated for the remainder of the 4-week experimental period. The administration of GTE or MGTE in high fructose-fed rats reduced postprandial blood TG significantly (P < 0.05). However, there was no significant difference in the postprandial blood TG responses to the GTE or MGTE treatment.
Table 2 summarizes the effect of GTE and MGTE treatment on fasting serum TC, HDL-C, NEFA, and uric acid concentrations. High fructose-fed rats had higher fasting serum TC levels (P < 0.05) than normal control diet-fed rats. GTE and MGTE administration in high fructose-fed rats decreased TC levels over the 4-week treatment period. However, there was no significant difference between the GTE and MGTE treated groups.
The HDL-C level was higher in the high fructose fed rats and decreased after GTE or MGTE treatment. Serum NEFA levels were significantly higher in the normal diet fed and the high fructose fed groups than in the HGTE and MGTE groups. GTE treatment reduced blood NEFA concentrations and MGTE treated rats had the lowest level of NEFA.
Enzyme activities
The activities of ME, which regulates lipogenesis in the liver and fat tissue, are presented in Table 3. The activity of ME in the liver was significantly greater in rats fed a high fructose diet than in rats fed a normal control diet. The activities of ME in the liver (NCON group, 53.4 ± 4.8; HCON group, 94.6 ± 5.1; HGTE group, 101.3 ± 3.4; HMGT group, 102.8 ± 4.0 mU/mg protein) did not differ among high fructose-fed groups irrespective of treatment with GTE. The activities of ME in the retroperitoneal fat tissue did not differ among the experimental groups.
The activity of catalase, which reduces the oxidative status, was significantly decreased in high fructose-fed rats (Table 3). The administration of GTE or MGTE increased the activity of catalase. The activity of hepatic catalase in the MGTE treated group was significantly higher than that in the GTE treated group. Renal catalase activity was not significantly different between the experimental groups, although a tendency towards a reduction of catalase activity was observed in the high fructose-fed groups.
Cardiac histopathology
Myocardial fibrosis was observed in the heart as shown in Table 4 and Fig. 2. The high fructose diets significantly increased the fibrosis compared to normal control diets (P < 0.001). The administration of MGTE, but not GTE, significantly decreased myocardial fibrosis in HGTE rats (Table 4 and Fig. 2). The presence of myocardial fibrosis in the HMGT group was reduced to the level in normal control rats. The mean LV thickness and LV luminal diameters were not significantly different among the experimental groups.
Discussion
This study examined the effect of non-encapsulated and microencapsulated green tea extract in rats fed a high fructose diet. Feeding high fructose for 8 weeks induced the development of several features of MetS. First, in rats fed a high fructose diet, serum TG, TC, and uric acid levels increased compared to that in those given normal control diet. These data are in agreement with several previous reports [32-34]. Bantle et al. [35] suggested that the effect of dietary fructose on blood TG is greater in the postprandial state than in the fasting state. In our study, postprandial blood TG increased > 100% in rats fed a high fructose diet compared to rats fed normal control diet. Hypertriglyceridemia due to high fructose intake has been attributed to an overproduction of lipid in the liver. The enhanced hepatic TG was related to overproduction of TG and decreased TG clearance [36]. In addition, the increased intake of fructose has been reported to enhance oxidative damage in rats [37]. In the present study, the high fructose diet might have increased the level of oxidative stress by decreasing catalase activities in the liver and kidney. Second, high fructose diet increased blood TC concentrations, body weight gain, and retroperitoneal fat mass.
Administration of 50 mg·kg·bw-1·d-1 GTE to rats with hypertriglyceridemia lowered the level of serum TG. Beneficial changes in serum TC levels were observed in both GTE and MGTE treated groups. GTE treatment decreased serum NEFA levels, resulting in normalization of serum TG (Table 2). The hypertriglyceridemia reported previously is likely related to increased de novo lipogenesis in combination with an increased NEFA flux into the liver [38,39]. Because the NEFA of adipocytes drain into the portal vein, they are delivered to the liver thereby stimulating gluconeogenesis, increasing TG synthesis, and ultimately resulting in the development of dyslipidemia [39]. GTE treatment decreased visceral obesity by decreasing the weight of visceral fat pad (epididymal and retroperitoneal fat pad). In addition, GTE treatment might be responsible for the observed reduction of serum NEFA levels in the HGTE and MGTE groups (Tables 1 and 2). In a previous study, catechin-rich GTE had an anti-obesity effect by elevating the metabolic rate and fat oxidation through lipolysis in adipose tissue [28]. In our study, MGTE treatment was associated with the lowest level of serum NEFA among all the experimental groups. However, there was no significant difference between serum TG and TC between the HGTE and MGTE groups. The malic enzyme, one of the lipogenic enzymes of liver and retroperitoneal tissue, was not affected by treatment with GTE or MGTE. This finding suggests that GTE might have a suppressive effect on hypertriglyceridemia by increasing lipid oxidation, rather than by decreasing the lipogenic enzyme activity. Unexpectedly, we observed an increase in HDL cholesterol in the HCON, HGTE, and MGTE groups. Elevated HDL-C levels are probably caused by increased cholesterol efflux from the peripheral and/or liver cells.
There is supporting evidence that uric acid may have a pathogenic role in the development of MetS. Hyperuricemia has been reported to predict the development of obesity and hypertriglyceridemia [25]. Nakagawa et al. [25] and other study reported that lowering the levels of uric acid improved some features of the MetS including hypertension, obesity, and hypertriglyceridemia [40]. In the present study, GTE prevented and reversed the features of MetS in fructose-fed rats. Microencapsulated GTE lowered the level of serum uric acid to a greater extent than GTE. It has been suggested that increased serum uric acid might induce the overproduction of hepatic TG and reduction of TG clearance through the decrease in lipoprotein lipase activity in endothelial cells [25]. In addition, uric acid could play a pathogenic role in fructose-mediated MetS by reducing endothelial NO bioavailability both in cell culture and in experimental models [40].
The present study provides additional information on the damaging effect of high fructose diet on the cardiovascular system by increasing myocardial fibrosis. Myocardial fibrosis, LV thickness, and LV diameter characterize the morphologic changes in rats and are gross indicators of cardiomyopathy [41]. Myocardial fibrosis is one of the main structural pathologic changes that develop over time in normal and obese animals [42], and has been associated with concurrent decrease in left ventricular compliance and varying degrees of ventricular hypertrophy [41]. Because the contribution of cardiovascular disease to human morbidity and mortality is very high in the elderly, much research in this area has been directed towards determining the role of specific food components on the development, progression, and prevention of CVD [4-9]. Our results demonstrate that MGTE treatment effectively prevents myocardial fibrosis together with a reduction in blood TG and TC levels. Recently, we demonstrated that microencapsulated GTE reduced the inflammatory response by decreasing NO production in murine RAW264.7 cells [43]. It is generally believed that catechin derivatives are the active components responsible for the beneficial effects of antioxidant [13] and cardioprotectants [44]. Oxidants are involved in the pathogenesis of dyslipidemia and CVD [12]. Therefore, the beneficial effects of antioxidants may be attributed, in part, to the lowering of oxidant levels. However, bioactive ingredients including polyphenols are degraded during processing and storage (temperature, oxygen, and light) and under the conditions encountered in the gastrointestinal tract (pH, enzyme, and interactions with other nutrient) [16]. The effectiveness of nutraceutical products in preventing various diseases depends on preserving the bioavailability of the bioactive ingredients. It has been reported that only a small proportion of the ingredients remain available following oral administration due to insufficient gastric residence time, low permeability, and/or solubility within the gastrointestinal tract [45]. In addition, Lambert et al. [46] found that polyphenols in green tea oxidize readily in beverages and even in the oral cavity, generating substantial levels of H2O2, which is potentially cytotoxic. Some bioactives are particularly susceptible to oxidation; therefore, microencapsulation of bioactives has the ability to preserve them in certain environments [47]. Lee et al. [48] found that microencapsulation of GTE with maltodextrin increased DPPH radical scavenging activities. Reineccius et al. [49] enhanced the retention of orange oil and prevented the oxidation by using gum Arabic. In our experiments, the beneficial effect of microencapsulated GTE was demonstrated by the reduction of serum uric acid and myocardial fibrosis and the increase in liver catalase activity compared to non-encapsulated GTE. Thus, the protection of bioactives from oxidation could be an effective way to utilize their beneficial effects on human health.
This work compared the ability of GTE and microencapsulated GTE to ameliorate hypertriglyceridemia and mediate cardiovascular protection. According to our results, microencapsulated GTE is more effective at alleviating the cardiovascular risk induced by high fructose feeding than non-encapsulated GTE.





 XML Download
XML Download