

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Type II diabetes is usually caused by peripheral insulin resistance and includes severe metabolic defects in glucose and lactate homeostasis such as hyperglycemia and hyperlactatemia [1,2]. These conditions are caused by an accumulation of glucose, insulin, and lactic acid in the blood, which causes the body to become more insulin resistant or excessively acidic.
Several lines of evidence indicate that resting blood glucose and lactate levels are elevated in diabetic rats [3-5]. In particular, several studies have reported that glucose and lactate expression in muscle tissue of diabetic rats is remarkably reduced [4-6]. Thus, one of the treatment options for insulin resistant diabetes, including hyperglycemia and hyperlactatemia, is to elevate glucose and lactate metabolic protein expression in skeletal muscle.
We have evaluated interventions that improve glucose and lactate metabolism in blood and muscles of diabetic animals including diet, physical exercise, and the use of pharmaceutical and nutriceutical compounds. Recently, epidemiological studies have shown a protective association between exercise training or dietary antioxidants and the development of type II diabetes [6-8].
Exercise training or an active life style have a profound benefit on glucose and lactate pathogenic phenotypes of insulin-resistant states. Indeed, several studies have shown that a transition from a sedentary life-style to a simple active lifestyle may be sufficient to develop favorable glucose and lactate metabolic adaptations and ameliorate pathogenic phenotypes in patients and animals with diabetes [4,9]. Therefore, exercise training has been included in diabetic treatment plans.
Increasing evidence indicates that diabetes may be associated with increased oxidative stress, and that antioxidant treatment may be beneficial. Selenium is a trace element that improves the antioxidant system by enhancing glutathione levels [10] and may exert beneficial effects on glucose metabolism, possibly via insulin-like actions, which may delay diabetes complications [7,11,12]. Glutathione peroxidase activity depends on selenium supplementation [13]. High doses of selenium are associated with memory loss, muscle pain, diarrhea,, and vomiting [14], whereas low selenium intake is associated with impaired glucose tolerance and gestational hyperglycemia [15]. According to the recommended daily allowance and the World Health Organization, optimum selenium intake is 50-200 µg/day.
At present, it is unknown whether selenium treatment affects hyperlactatemia or if lactate levels change in the blood and muscles of diabetic rats following selenium treatment. Although it is clear that exercise training alone or selenium alone effectively ameliorates some pathogenic blood and muscle phenotypes in diabetes, there is currently no information regarding how these two interventions might interact when applied in combination to ameliorate glucose and lactate blood and muscle pathogenic phenotypes in a diabetic animal model that expresses human-like phenotypes.
Therefore, we conducted this study to determine if exercise training and selenium treatment in combination improves glucose and lactate pathogenic phenotypes of diabetes using Goto-Kakisaki (GK) rats, which are a highly inbred strain of Wistar rats that spontaneously develop type II diabetes without obesity but exhibit insulin resistance and hyperglycemia and resemble the polygenic basis for disease in the majority of patients with type 2 diabetes [16].
Specifically, GK rats underwent a 6 wk treatment with exercise training and selenium individually or in combination, after which the levels of blood metabolic parameters and a number of proteins involved with glucose metabolism, such as AMP-activated protein kinase (AMPK), peroxisome proliferator-activated receptor coactivator 1α (PGC-1α), and glucose transporter 4 (GLUT-4), and lactate metabolism, such as lactate dehydrogenase (LDH), monocarboxylate transporter 1 and 4 (MCT1 and 4), and cytochrome oxidase IV (COX-IV) in skeletal muscles were determined. The results are particularly important in the context of designing interventions that substantially ameliorate glucose and lactate pathogenic phenotypes of diabetes.
Materials and Methods
Animals and treatments
All experimental procedures were approved by the Committee on Animal Research at Korea National Sport University. Eight week old male GK rats (n = 40; mean weight, 280.30 ± 20.35 g) were obtained from Charles River (Yokohama, Japan) and cared for according to the Guiding Principles for the Care and Use of Animals, based on the Helsinki Declaration of 1964 (American Physiology Society Operational Guide Revision, 2002) [17]. The rats were studied from 46 weeks of age (n = 26, mean weight, 336.05 ± 4.13 g) to 52 weeks of age (n = 26, mean weight, 365.16 ± 38.23 g).
Rats were maintained on a 12:12 hour dark-light cycle, housed at 22 ± 2℃ with a 50% relative humidity, and had free access to a standard chow diet ad libitum. The body weights of the animals were measured at the start of the study and after the training period in each group. GK rats were randomly assigned to one of the following groups: a sedentary control group (SED), an exercise trained group (ET), selenium treated group (SEL), or a combined selenium treated and exercise trained group (COMBI).
Sodium selenite (S5261, Sigma, St. Louis, MO, USA) was dissolved in distilled water to a final concentration of 0.2 µmol/µl. Animals in the SEL groups received 5 µmol/kg body weight/day of purified sodium selenite, dissolved in 0.2 ml dH2O by intraperitoneal injection every morning for 6 weeks, as described previously [18]. The SED animals received the same dose of dH2O. Animals in the ET group ran 50 minutes per day at 21 m/min 5 days/week at a 0% gradient for 6 weeks. Prior to the treadmill ET, the rats were allowed a minimum 2 week adaptation period to the new environment, consisting of running for 10 minutes a day on the treadmill (11 m/min). The COMBI animals performed the treadmill training protocol exactly as described above, and also received the daily selenium treatment.
Intraperitoneal glucose tolerance test (IPGTT)
Twenty-four hours after the final ET bout or selenium treatment, the four groups underwent an IPGTT, as previously described by Hamamoto et al. [19]. Rats were intraperitoneally administered a glucose solution (1 g/kg body weight in 0.9% NaCl). Blood was withdrawn from a cut at the tip of the tail at 0, 30, 60, 90, and 120 min after glucose administration to measure plasma glucose. Blood glucose levels were determined by the sensitive strip method using a blood glucose monitoring system (I-sens Co., Seoul, South Korea), and the homeostasis model of assessment (HOMA-IR, µIu/ml × mmol/L/22.5) was calculated. This latter variable is defined as an index of insulin resistance, and reduced values reflect an increase in whole-body insulin sensitivity.
Tissue preparation
Cardiac and skeletal muscles were excised, weighed, cleaned of any visible blood, frozen in liquid nitrogen, and stored until further biochemical analysis.
Glycogen content
Muscles and liver glycogen content was assayed as previously described by Lowry and Passonneau [20]. Briefly, 10 mg of soleus and plantaris muscles as well as liver samples were placed in 500 µl of 2 N HCl (Duksan Pure Chemical Co., Ltd., Kyungki-Do, Korea), and incubated at 100℃ for 2 hr. After neutralization with 1.5 ml of 0.66 N NaOH ( D-6100; Merck, Darnstadt, Germany), the liberated glucose units were assayed fluorometrically (Genios, TECAN, Vienna, Austria), and glycogen content was expressed as micromoles of glucosyl units liberated per gram of wet muscles and liver weight (µmol·g-1 w·w.).
Citrate synthase (CS)
To monitor muscle adaptation to endurance training, the soleus and plantaris muscles as well as the heart were assayed using a standard spectrophotometer method for maximal CS activity after 6 weeks of training in the ET rats and in SED rats. We determined CS activity (units g-1 tissue) in triplicate at 30℃ for each muscle using methods modified from Metz et al. [5]. Fresh homogenates, kept on ice, were used for assays. Frozen tissues (5-10 mg) were homogenized on ice using 40 volumes of extraction buffer (pH 7.5). consisting of 50 mM Tris/HCl, 5 mM MgSO4, 1 mM ethylene diamine tetra-acetic acid, 1 mM reduced glutathione, and 0.1% Triton X-100. CS activity was estimated as the reduction of 5,5 dithiobis-(2-nitrobenzoic) acid (DTNB) at 412 nm over 2.5 min using a microplate reader (Model 550; Bio-Rad, Tokyo, Japan). Results are expressed in micromoles per minute per milligram of protein. Aliquots of original homogenates were further diluted by 300 for CS samples. The CS assay medium consisted of 100 mM Tris/HCl (pH 8.0), 1.0 mM DTNB, 3.0 mM acetyl CoA and 5.0 mM oxaloacetate. The mM extinction coefficient of DTNB is 14.1.
Isolation of mitochondria
Mitochondria isolated from myocardium were extracted using a mitochondria extraction kit (IMGENEX Corp., San Diego, CA, USA) as follows: 100 mg of each sample was homogenized in 1 ml homogenizing buffer and centrifuged at 3,000 rpm for 10 min at 4℃, and the supernatant was removed. The resulting supernatant was centrifuged again at 12,000 rpm for 30 min at 4℃. The pellet was mixed with 1 ml of suspension buffer and then centrifuged at 12,000 rpm for 10 min at 4℃. The resulting pellet was mixed with 1 ml of suspension buffer for 30 min at 4℃. The supernatant was recovered as the mitochondrial fraction.
Western blot analysis
Proteins (80 µg/lane) prepared from the tissues of diabetic GK rats were separated for 90 min using electrophoresis on a 10% SDS/PAGE gel and then transferred to polyvinylidene membranes for 1 hr at 60 volts. Each membrane was separately incubated overnight at 4℃ with one of the following primary antibodies: anti-AMPK (#-2532) and p-AMPK (#-2531) antibodies (Cell Signaling Technology, Danvers, MA, USA, 1:1000). PGC-1α (sc-13067), GLUT-4 (sc-1608), LDH (sc-33781), MCT-1 (sc-14917), COX-IV (sc-8385), GAPDH (sc-20357) antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA; 1:1000), and MCT-4 (AB 3314P) antibody (Chemicon, Temecula, CA, USA;1:500). The membranes were washed with washing buffer (150 mM NaCl, 10 mM Tris-HCl, 0.05% Tween 20, pH 8.0) and then incubated with secondary antibodies: horseradish peroxidase (HRP)-conjugated donkey anti-goat (Santa Cruz Biotechnology, sc-2004; 1:5000) for GLUT-4, MCT-1, mtMCT-1, and GAPDH; HRP-conjugated goat anti-rabbit (Santa Cruz Biotechnology, sc-2020; 1:5000) for PGC-1α, LDH, mtLDH, AMPK, p-AMPK, and MCT-4; and HRP-conjugated goat anti-mouse (Santa Cruz Biotechnology, sc-2005; 1:5000) for COX-IV. The membrane blot was detected with an enhanced chemiluminescence Western blot analysis system (Santa Cruz Biotechnology), following the manufacturer's instructions. The density of the developed bands was scanned using a ChemiDoc XRS system (Bio-Rad, Hercules, CA, USA).
Biochemical analysis
Whole blood was extracted precisely 24 hours after the final ET or selenium treatment by cardiac puncture using an 18-gauge needle with a 10 ml syringe. After extraction, blood samples were centrifuged at 15,000 rpm, at 4℃ for 10 min, and the supernatants were transferred to new tubes and stored at -80℃ until lactate (Lactate kit, Roche Diagnostics, Indianapolis, IN, USA), glucose (Glucose Hexokinase kit, Bayer, Pittsburgh, PA, USA) and insulin (Insulin RIA kit, Linco Research Inc, St. Charles, MO, USA) were analyzed.
Statistical analysis
Data were analyzed using SPSS version 18.0 (SPSS, Inc., Chicago, IL, USA). All values are expressed as means ± SEs. A two-way mixed analysis of variance (ANOVA) was used to evaluate statistical significance of the IPGTT. Statistical significance was determined using a one-way ANOVA when comparing the groups. A Bonferroni post hoc test was followed for all pairwise multiple comparisons if a statistically significant main effect was found. Differences were considered statistically significant at α = 0.05.
Results
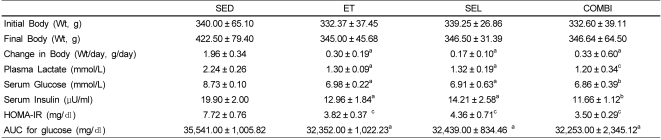
Effect of ET and SEL on body weight and metabolic parameters
A one-way ANOVA on body weight [F(3,20) = 12.74, P < 0.001], lactate, [F(3,20) = 4.08, P < 0.05], glucose [F(3,20) = 4.88, P < 0.01], insulin [F(3,20) = 3.42, P < 0.05], HOMA-IR [F(3,20) = 4.15, P < 0.05], and glucose area under curve (AUC) [F(3,20) = 6.20, P < 0.05] data indicated significant effects by group (Table 1). The ET, SEL, and COMBI groups had significantly lower final body weights, and lactate, glucose, and insulin levels, as well as HOMA-IR and glucose AUC relative to the SED (Table 1).
Effect of ET and SEL on glucose response
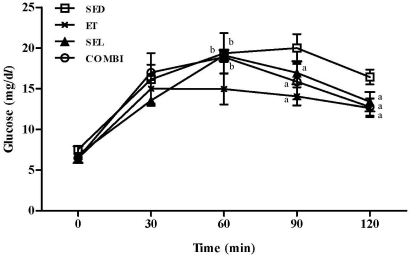
According to a two-way mixed ANOVA, an interaction was observed between groups and times (0, 30, 60, 90, and 120 min) [F(4,12) = 8.29, P < 0.001]. A statistically significant difference over time was observed. A one-way ANOVA on glucose response on the 60 min [F(3,20) = 7.66, P < 0.01], 90 min [F(3,20) = 9.20, P < 0.01], and 120 min [F(3,20) = 11.75, P < 0.001] data indicated significant group effects (Fig. 1). ET resulted in a lower glucose response at 60 min (P < 0.05) relative to the other groups, whereas ET, SEL, or COMBI resulted in a reduced glucose response at 90 (P < 0.05, respectively) and 120 min (P < 0.05, respectively) relative to the SED (Fig. 1).
Effect of ET and SEL on glycogen content and CS activity in skeletal muscles and liver
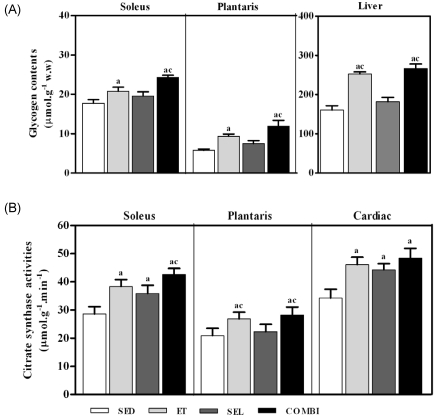
A one-way ANOVA on glycogen content in the soleus [F(3,20) = 11.85, P < 0.01] and plantaris [F(3,20) = 29.46, P < 0.001] muscles and liver [F(3,20) = 76.98, P < 0.001] data indicated significant group effects (Fig. 2A). The glycogen content was higher in the soleus and plantaris muscles and liver of the ET and COMBI groups (P < 0.05, vs. SED, respectively). COMBI resulted in a significantly higher glycogen content in the soleus and plantaris muscles and liver (P < 0.05, vs. SEL, respectively). Moreover, ET resulted in a significant increase in glycogen content in the liver (P < 0.05, vs. SEL). A one-way ANOVA on CS activity in the soleus [F(3,20) = 82.22, P < 0.05] and plantaris [F(3,20) = 35.46, P < 0.05] muscle and cardiac [F(3,20) = 34.51, P < 0.001] data indicated significant group effects. ET, SEL, or COMBI resulted in significant increases in CS activity in the soleus (P < 0.05 vs. SED, respectively) and cardiac muscles (P < 0.05, respectively, vs. SED) (Fig. 2B). Additionally, COMBI resulted in a significant increase in CS activity in the soleus (P < 0.05 vs. SEL). Finally, ET and COMBI induced significant increases in CS activity in the plantaris muscle (P < 0.05, vs. SED, respectively; P < 0.05, vs. SEL, respectively) (Fig. 2).
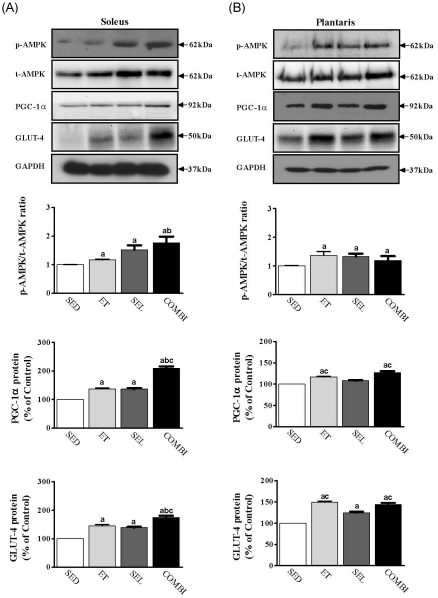
Effect of ET and SEL on AMPK phosphorylation, PGC-1α, and GLUT-4 proteins in the soleus and plantaris muscles
A one-way ANOVA on pAMPK/AMPK [F(3,20) = 18.36, P < 0.001], PGC-1α [F(3,20) = 29.15, P < 0.001], and GLUT-4 [F(3,20) = 1605.72, P < 0.001] (Fig. 3A) in soleus muscle indicated significant group effects. The levels of AMPK phosphorylation, and PGC-1α, and GLUT-4 protein in the soleus muscle in the ET, SEL, and COMBI were higher than those in the SED group (P < 0.05, respectively). Furthermore, COMBI had significantly greater levels of AMPK phosphorylation and PGC-1α and GLUT-4 protein content in the soleus muscle relative to ET (P < 0.05, respectively). Moreover, PGC-1α and GLUT-4 protein levels in the soleus muscle in the COMBI were higher than those in the SEL (P < 0.05, respectively). A one-way ANOVA on pAMPK/AMPK [F(3,20) = 5.65, P < 0.05], PGC-1α [F(3,20) = 29.15, P < 0.001], and GLUT-4 [F(3,20) = 173.77, P < 0.001] in plantaris muscle indicated significant group effects. ET, SEL, and COMBI resulted in a significant increase in AMPK phosphorylation and GLUT-4 protein levels in the plantaris muscle (P < 0.05, vs. SED, respectively) (Fig. 3B). Additionally, ET and COMBI had significantly higher PGC-1α and GLUT-4 protein levels in the plantaris muscle relative to the SEL (P < 0.05, respectively). Finally, ET and COMBI had a significantly higher PGC-1α protein level in the plantaris muscle relative to the SED (P < 0.05, respectively) (Fig. 3).
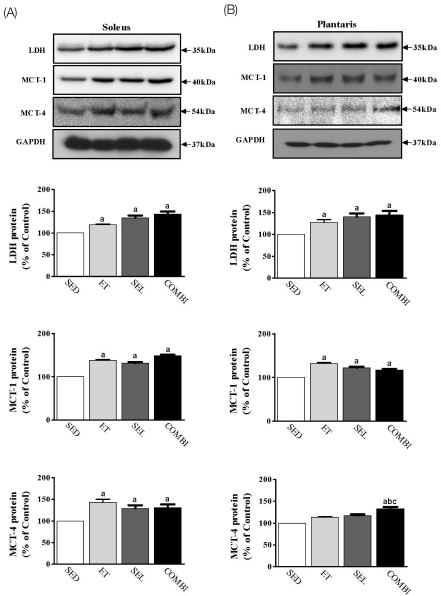
Effect of ET and SEL on LDH, MCT-1, and MCT-4 proteins in the soleus and plantaris muscles
A one-way ANOVA on LDH [F(3,20) = 19.74, P < 0.001], MCT-1 [F(3,20) = 498.90, P < 0.001], and MCT-4 [F(3,20) = 14.72, P < 0.001] in the soleus muscle indicated significant group effects (Fig. 4A). ET, SEL, and COMBI resulted in significant increases in the LDH, MCT-1, and MCT-4 protein levels in the soleus muscle (P < 0.05, respectively, vs. SED). A one-way ANOVA on LDH [F(3,20) = 8.21, P < 0.01], MCT-1 [F(3,20) = 52.24, P < 0.001], and MCT-4 [F(3,20) = 27.61, P < 0.001] in plantaris muscle indicated significant group effects (Fig. 4B). ET, SEL, and COMBI resulted in significant increases in LDH and MCT-1 protein levels in plantaris muscle (P < 0.05, vs. SED, respectively). COMBI exerted a significant increase in the MCT-4 protein level in plantaris muscle (P < 0.05, vs. SED; P < 0.05, vs. ET; P < 0.05, vs. SEL, respectively) (Fig. 4).
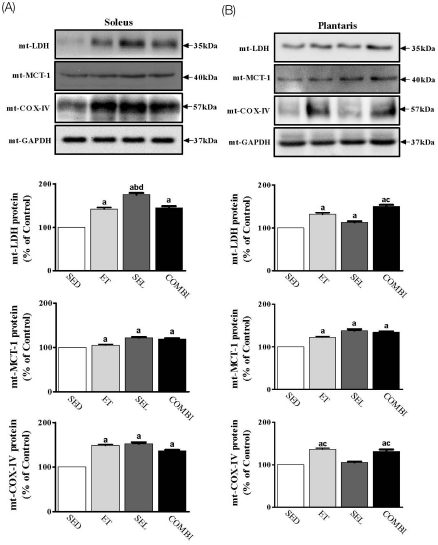
Effect of ET and SEL on mitochondrial (mt) LDH, MCT-1, and COX-IV proteins in soleus and plantaris muscles
A one-way ANOVA on mtLDH [F(3,20) = 839.47, P < 0.001], mtMCT-1 [F(3,20) = 35.83, P < 0.001], and mtCOX-IV [F(3,20) = 713.33, P < 0.001] in soleus muscle indicated significant group effects (Fig. 5A). ET, SEL, and COMBI resulted in significant increases in mtLDH, mtMCT-1, and mtCOX-IV protein levels in soleus muscle (P < 0.05, vs. SED, respectively). SEL induced a significant increase in the mtLDH protein level in the soleus muscle (P < 0.05, vs. ET; P < 0.05, vs. COMBI, respectively). A one-way ANOVA on mtLDH [F(3,20) = 144.31, P < 0.001], mtMCT-1 [F(3,20) = 109.47, P < 0.001], and mtCOX-IV [F(3,20) = 110.35, P < 0.001] in plantaris muscle indicated significant group effects (Fig. 5B). ET, SEL, and COMBI led to significant increases in the mtLDH and mtMCT-1 protein levels in the plantaris muscle (P < 0.05, vs. SED, respectively). COMBI resulted in a significant increase in the mtLDH protein level in the plantaris muscle (P < 0.05, vs. SEL, respectively). ET and COMBI resulted in a significant increase in the mtCOX-IV protein level in plantaris muscle (P < 0.05, vs. SED; P < 0.05, vs.SEL, respectively) (Fig. 5).
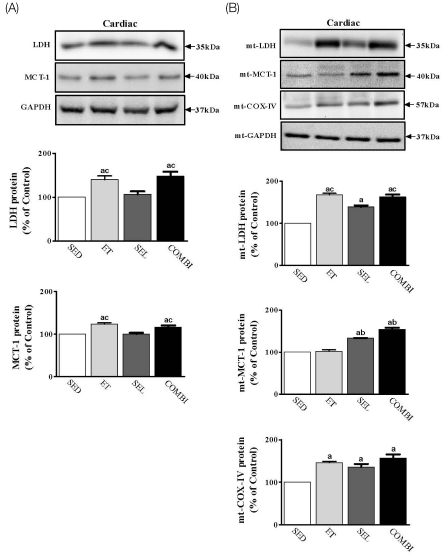
Effect of ET and SEL on total or mtLDH, MCT-1, and COX-IV proteins in cardiac muscle
A one-way ANOVA on LDH [F(3,20) = 10.12, P < 0.01], mtLDH [F(3,20) = 448.08, P < 0.001], MCT-1 [F(3,20) = 24.97, P < 0.001], mtMCT-1 [F(3,20) = 136.00, P < 0.001], and mtCOX-IV [F(3,20) = 47.68, P < 0.001] in cardiac muscle indicated significant group effects (Fig. 6A). ET and COMBI resulted in significant increases in LDH and MCT-1 protein levels in cardiac muscle (P < 0.05, vs. SED; P < 0.05, vs. SEL, respectively. ET, SEL, and COMBI resulted in significant increases in mtLDH and mtCOX-IV protein levels in cardiac muscle (P < 0.05, vs. SED, respectively). ET and COMBI produced a significant increase in the mtLDH protein level in cardiac muscle (P < 0.05, vs. SEL, respectively). SEL and COMBI resulted in a significant increase in the mtMCT-1 protein level in cardiac muscle (P < 0.05, vs. SED; P < 0.05, vs. ET, respectively) (Fig. 6).
Discussion
The benefits of ET and antioxidants are well established but are seldom available to people suffering from diabetes. Although it may not be possible to prevent the underlying disease process, the benefits of ET and selenium for managing patients with diabetes have been demonstrated. ET is regarded as a useful strategy to ameliorate hyperglycemia, hyperlactatemia, and insulin resistance in patients with diabetes [21,22,34]. Moreover, selenium is an essential micronutrient that improves glucose tolerance [15] and reduces oxidative stress [10] and insulin resistance in mammals [7].
However, despite growing evidence of the benefits of ET and SEL for the health and function of patients with diabetes, little clinical evidence supports the recommendations for ET guidelines and SEL supplementation in patients with diabetes. Therefore, we evaluated the protective effects of ET and SEL individually and in combination on molecular pathogenic phenotypes associated with type II diabetes.
We demonstrated that ET increased muscle and liver glycogen content and CS activity in diabetic GK rats, while reducing measurable levels of plasma lactate, serum glucose, insulin, HOMA-IR, and the glucose AUC. These results suggest that ET ameliorates impaired glucose metabolism and insulin resistance likely occurring via muscle stimulated-insulin action to improve glucose tolerance, increase enzyme activity involved in glucose oxidation (CS), and initiate the induction of metabolic genes associated with aerobic glucose and lactate metabolism in diabetic animals. These findings are in agreement with previous studies [4,5,21,22].
Mckenzie et al. [11] reported that excessive amounts of SEL cause selenosis, whereas a medium dose (55 µg/day) is recommended for health. In our report, SEL administration in GK rats was 5 µmol/kg body weight/day, which is similar to that reported to be effective for alleviating diabetic symptoms in diabetic rats [18,23] and its effects on glucose and lactate metabolism were then evaluated. It is clear from the present findings that SEL alone reduced blood lactate, glucose, and insulin levels along with an increase in muscle CS activity, which could have led to increased skeletal muscle lactate clearance, similar to the results of Fürnsinn et al. [24] and Hwang et al. [18], who suggested that SEL alone may exert either beneficial extra-pancreatic action on hypoglycemic and hypolactatemic effects in diabetic animals. However, our results also suggest that SEL alone did not increase liver or muscle glycogen content compared to those in SED animals.
COMBI reduced blood lactate, glucose, insulin levels, HOMA-IR, and glucose AUC as well as elevated glycogen content and CS activity in liver and muscles. These results suggest that ET leads to improved glucose homeostasis and oxidative metabolism and ameliorates insulin resistance in sedentary diabetic GK rats, which leads to a selenium treatment-induced insulin-like improvement in glucose homeostasis and oxidative metabolism similar to that reported in diabetic rats that have undergone ET [25,26]. However, we found no evidence that COMBI of diabetic GK rats beneficially modified either muscle glycogen content or CS activity relative to ET or SEL alone.
One important finding of this study was that ET alone, SEL alone, or the COMBI regimen was effective for significantly enhancing AMPK, PGC-1α, and GLUT-4 protein levels in skeletal muscle of diabetic GK rats. These changes were induced by muscle-stimulated insulin action [27,28] and insulin-like action on supplemented SEL [21,22] or both with a clinical reduction in blood metabolic parameters.
However, the expression of AMPK, PGC-1α, and GLUT-4 proteins in the ET alone, SEL alone, or COMBI appeared to be tissue-specific and not what we expected. The COMBI did not result in an additive effect of ET and SEL on AMPK, PGC-1α, and GLUT-4 expression in skeletal muscle relative to ET or SEL alone. Nevertheless, it is clear that muscle-stimulated insulin action and SEL-treated insulin-like action or both increased muscle AMPK and GLUT-4 protein levels, possibly via up-regulation of the PGC-1α protein in resistant skeletal muscles.
However, these results have limitations because we did not measure GLUT-4 membrane localization or glucose uptake, and we did not measure AMPK- or PGC-1α-associated GLUT-4 promoter activity when applied to ET alone, SEL alone, or COMBI. Therefore, further research is needed to identify the aforementioned variables in diabetic GK rats, possibly through mechanisms underlying the beneficial effect of ET alone, SEL alone, or COMBI.
Our results also showed that ET and SEL increased lactate metabolic proteins (LDH, MCT-1, MCT-4, COX-IV) in diabetic GK rats, which was associated with an increase in insulin sensitivity and oxidative enzymatic activities. There is evidence that elevated lactate levels lead to a decrease in muscle lactate transport capacity by down regulating known lactate metabolic proteins and CS activity in insulin-resistant states [4,5,29]. Therefore, we examined the effects of ET alone, SEL alone, or COMBI on the expression of proteins (LDH, MCT-1, MCT-4, COX-IV) involved in lactate metabolism in diabetic GK rats. As a result, ET increased total or mtLDH, MCT-1, and COX-IV proteins along with CS activity in skeletal and cardiac muscle, similar to the results reported by McCullagh et al. [30], Hughes et al. [31], Enoki et al. [4], and Metz et al. [5] who suggested that LDH or MCT-1, oxidative capacity, and lactate oxidation are strongly correlated and that upregulation of LDH, MCT-1, and COX-IV proteins reduces hyperlactatemia in a diabetic animal model after ET.
In general, tissue specificity of MCT-4 expression is highly related to muscle glycolytic capacity [32]. However, our results showed that ET increased the MCT-4 protein only in the soleus, which was inconsistent with the results of studies conducted by Bonen et al. [32] and Juel and Halestrap [33], who suggested that MCT-4 is related to glycolytic muscle fibers. Therefore, further research should identify tissue-specific MCT activity in diabetic GK rats, possibly via ET.
The role of SEL treatment on lactate metabolic proteins has not been previously evaluated in diabetic animals. We have provided new information regarding the potential utility of SEL treatment for increasing LDH, MCT-1, MCT-4, and COX-IV proteins in insulin-resistant muscle. A significant finding was that SEL treatment increased total or mtLDH and MCT-1 and mt-COX-IV proteins, reduced lactate level, but elevated CS activity in skeletal muscle relative to those in SED. In addition, SEL increased the mtLDH and mtMCT-1 proteins but not LDH and MCT-1 in cardiac muscle. Therefore, the SEL effect on total or mtLDH, MCT-1, and COX-IV expression appears to be tissue-specific. Additionally, SEL also increased the MCT-4 protein in soleus muscle only, an adaptive process similar to that observed with ET in diabetic GK rats. Hence, upregulation of total or mtLDH, MCT-1, and MCT-4 in skeletal muscle can be regulated by supplemented selenium through an insulin-like action and a decrease in lactate, glucose, and insulin levels, suggesting that SEL reduces lactate level, which could be more related to an increase in lactate uptake.
However, the mechanism by which SEL treatment induces an upregulation of lactate metabolic proteins in sedentary diabetic GK rats was not determined because no lactate fluxes across the sarcolemma or MCTs translocation were observed in either skeletal or cardiac muscle in the present study. Therefore, additional experiments are needed to understand the underlying mechanisms involved in the increased muscle lactate uptake that occurred in diabetic GK rats.
We found that COMBI induced increases in LDH, MCT-1, MCT-4, and COX-IV proteins in skeletal and cardiac muscles in diabetic GK rats, indicating that the upregulation of these proteins occurred in response to major changes in the specific activity of CS along with a significant reduction in the lactate level observed following the COMBI treatment.
Therefore, the underlying mechanism of the upregulation in total and mtLDH, MCT-1, and COX-IV proteins following the COMBI treatment may be linked to oxidative metabolism and strongly related to lactate oxidation in skeletal and cardiac muscles. Additionally, COMBI increased MCT-4 protein in both the soleus and plantaris muscle, suggesting that MCT-4 upregulation in these muscles may have contributed to lower lactate levels, but clearly other mechanisms must also come into play. Therefore, MCT-4 is regulated in different ways by COMBI depending on the tissue.
Expression of the glucose and lactate metabolic proteins was not further enhanced with COMBI compared to that of ET or SEL alone. Therefore, additional experiments are needed to understand the underlying mechanisms involved in the increased muscle lactate uptake that occurs in diabetic GK rats, because no fluxes in lactate across the sarcolemma or MCTs translocation were observed in either skeletal muscle or cardiac muscle in the present study.
In conclusion, ET alone, SEL alone, or COMBI showed a positive effect on ameliorating pathogenic phenotypes of diabetes possibly by upregulating AMPK-PGC-1α and GLUT-4 and lactate metabolic proteins, which appeared to be tissue specific. However, it appeared that COMBI did not have a synergistic effect. Taken together, these results suggest that ET alone, SEL alone, or COMBI is a simple behavioral and nutriceutical intervention sufficient to repress hyperglycemia and hyperlactatemia in animals with diabetes. However, further studies are needed to determine the interactive benefits of ET and SEL and the mechanisms underlying these beneficial effects.
 XML Download
XML Download