

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Hepatic fibrosis is a wound-healing response to chronic liver injury that leads to cirrhosis and liver failure if repetitive liver damage occurs. Exciting progress has been made in the understanding of the mechanisms of hepatic fibrosis, which demonstrates that the central event of liver fibrosis is the activation of stellate cells involving the transformation of quiescent vitamin-A-rich cells into proliferative, fibrogenic, and contractile myofibroblasts [1,2]. Therefore, targeting the stellate cells and fibrogenic mediators could be a mainstay of anti-fibrotic therapy [3]. Antioxidants are known to inhibit the activation of hepatic stellate cells (HSC) and expected to exhibit antifibrotic effects [4].
Grape, Vitis vinifera is one of the most popular and widely cultivated fruits in the world. Grape contains lots of active components, including flavonoids, polyphenols, anthocyanins, proanthocyanidins, procyanidins, and the stilbene derivatives resveratrol, and the major medicinal properties of grape and its constituents are anticarcinogenic, immunomodulatory, antidiabetes, anti-obesity, and anti-aging [5,6]. Although these bioactive components of grape mainly exist in grape skin and seeds, grape skin and seeds are usually discarded in regular dietary intake and the winery and grape juice industry. These wastes contain bioactive components with potent antioxidant and free radical scavenging activity. As reducing oxidative stress, which is an important stimulus to activation of hepatic stellate cells (HSC), is a relatively practical avenue of intervention [4], grape skin or seeds are expected to exhibit hepatoprotective and antifibrotic effects by reducing oxidative stress, which exerts protective effects against HSC activation.
Dimethylnitrosamine (DMN) is a potent hepatotoxin, carcinogen, and mutagen [7]. At doses as low as 20 mg/kg, DMN can cause massive liver necrosis and death. Exposure to repeated lower doses of DMN causes subacute and chronic liver injury with necrosis, fibrosis, and nodular regeneration [8]. DMN-induced fibrosis reproduces most of the features observed during human liver fibrosis [9]. This model shows progressive and obvious pathological alterations, high reproducibility of fibrosis, and low mortality rates in experimental animals [10]. Using the DMN-induced liver fibrosis model in rats, we investigated whether grape skin and seeds have an antifibrogenic effect.
Materials and Methods
Induction of liver fibrosis with DMN
Animal care and all experimental procedures were conducted in accordance with the Guide for Animal Experiments edited by the Korean Academy of Medical Sciences. Male Sprague-Dawley (SD) rats were obtained from Dae-Han Laboratory Animal Research Center Co., Ltd. (Seoul, Korea). Vitis vinifera was purchased at a common market, Korea. The grape skins were separated from the pulps, dried at 70℃ in a dry oven for several hours, and ground to powder using a grinder. The grape seed powder, which was dried at 50℃ and grinded to powder, was obtained from Pacific Food Industry (Gyeongbuk, Korea). The grape skin powder and seed powder were mixed uniformly with the basal diet powder at concentrations of 10% (w/w), respectively. Animals were fed ad libitum and had free access to both food in powder feeders and water in a temperature- and humidity-controlled animal house under 12-h light-dark cycles. The experimental diets of 6 groups are described in Table 1.
Thirty six rats, 140-160 g, were divided into 6 groups of 6 each: Normal, DMN, K10, DK10, S10, and DS10 groups. The DMN group was treated with intraperitoneal (i.p.) injections of DMN (diluted with saline) at 10 mg/kg per day for 3 consecutive days per week for 4 weeks [10,11]. The DK10 and DS10 groups were treated with DMN and fed daily a powdered diet mixed with 10% grape skin and seeds, respectively, for the length of the study. The K10 and S10 groups were fed the same diets alone. The normal group was treated with the volume of saline equivalent to the others groups. At the end of the fourth week, all rats were sacrificed under ether anesthesia and their livers were excised and weighed. Blood samples for biochemical analyses were obtained from the inferior vena cava. The liver specimens were immediately fixed in 10% neutral buffered formalin for histochemical studies. The remaining liver tissue was homogenized using a glass Potter-Elvehjem homogenizer set. The homogenate was freed from the cellular debris and nuclei by centrifugation at 700 ×g at 4℃ for 10 min. Protein concentration was determined by the method of Lowry et al. [12] using bovine serum albumin as the standard.
Biochemical analyses of serum
Serum aspartate transaminase (AST), alanine transaminase (ALT), and alkaline phosphatase (ALP) activities were estimated by colorimetric methods using commercial kits (Eiken, Tokyo, Japan). Serum albumin, total protein, and bilirubin levels were also measured using commercial kits following the manufacturer's protocols (Eiken, Tokyo, Japan).
Histology and immunohistochemistry
Five-micrometer liver sections were deparaffinized and processed routinely for hematoxylin-eosin (H&E) and Sirius red (SR) staining [13], and were examined immunohistochemically for α-smooth muscle actin (α-SMA, Serotec, Oxford, UK) using routine indirect avidin-biotin immunolabeling. Non-immune isotype-matched immunoglobulin was used in place of the primary antibody in the negative controls.
Image analysis
The mean values of SR or α-SMA positive areas were assessed in 6 ocular fields per specimen, which were randomly selected at 40x magnification using an image analysis system (Image Pro Plus 4.0, Media Cybernetics, Del Mar, CA, USA). The SR or α-SMA positive areas were determined as the mean of triplicate examinations and expressed as a percentage of the total area of the specimen.
Hydroxyproline and malondialdehyde (MDA) content determination
Hydroxyproline levels in the liver were determined according to the Woessner method [14]. Approximately 100 mg of liver tissue was hydrolyzed in 6 M HCl in a sealed tube at 110℃ for 24 h. The hydrolysates were evaporated and the residues were dissolved in distilled water and filtered. The filtrate was then mixed with 1.4% chloramine-T solution and perchloric acid. Finally, 20% p-dimethylaminobenzaldehyde in methylcellosolve was added, and placed in a water bath at 60℃ for 20 min. After cooling, absorbance was measured at 560 nm. Malondialdehyde levels were determined by the Buege and Aust method [15].
Results
Body and liver weight
The effects of the grape skin and seed on body and liver weight of rats are shown in Table 2. Treatment with DMN caused a significant decrease in both body weight and liver weight compared with the normal group. In the DMN group, body weight gain for 4 weeks was only 38% of the normal and supplementation of grape skin or grape seeds decreased this loss. In the DK10 and DS10 group, the body weight gains were 74% and 78% of the normal, respectively. In the DMN group, the liver weight was 63% of the normal. But supplementation of grape skin or seeds significantly prevented the DMN-induced loss of liver weight. There were no significant changes on body and liver weight among the K10 and S10 groups compared to the normal and there were no differences on the body weight and liver weight among the DK10 and DS10 groups.
Serum parameters of liver function
The effects of grape skin and seeds on serum parameters in the liver model are shown in Table 3. DMN-induced increases in serum AST, ALT and ALP levels were significantly suppressed by grape skin or seeds feeding. Again, in the DK10 and DS10 groups, diminished serum albumin and total protein concentrations were restored. Grape skin and seed treatment blocked increases in plasma bilirubin. These results indicate that grape skin and seeds protected against hepatocytes necrosis after DMN treatment. There were no changes on the serum parameters by feeding the grape skin and seeds only and there were no significant differences between the DK10 and DS10 groups on serum parameters.
Histopathology and immunohistochemistry
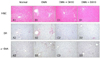
The effects of grape skin and seeds on DMN-induced liver injury were evaluated by histopathologic examination of the liver injury sections by H&E staining. In contrast to the normal group of rats (Fig. 1A1), the treatment of DMN for 4 weeks caused extensive hemorrhagic necrosis and disruption of tissue architecture (Fig. 1B1). These alterations were remarkably reduced in the liver sections of the grape skin-treated rats (Fig. 1C1) and the grape seed-treated rats (Fig. 1D1). Serial sections were stained with SR for collagen. In the liver sections taken from the normal, collagen fibers were observed only in the periportal area (Fig. 1A2). The livers of the DMN-treated rats exhibited an increase in collagen content, and displayed bundles of collagen fibers surrounding the lobules, forming large fibrous septa (Fig. 1B2). The thickening of these collagen fiber bundles was markedly reduced in the grape skin-treated group (Fig. 1C2) and in the grape seed-treated group (Fig. 1D2). Quantitative analysis by an image analysis technique also showed a remarkable reduction of SR staining regions in the grape skin-treated group and the grape seed-treated group, compared with the DMN group (Table 4). The expression of α-SMA, an indicator of activated HSCs, was detected by the immunohistochemistry method. In contrast to the normal group (Fig. 1A3), many α-SMA-positive cells were detected around the periportal fibrotic band areas and were scattered in the regions of connective tissue septa in the DMN-treated rats (Fig. 1B3). However, in the DK10 group and DS10 group, only traces of α-SMA-positive cells were detected (Fig. 1C3 and Fig. 1D3). Quantitative analysis by image analysis technique showed a remarkable reduction of α-SMA-positive regions in the grape skin-treated group and the grape seed-treated group, compared with the DMN group (Table 4).
Hepatic hydroxyproline
DMN caused a 1.79-fold increase in hepatic hydroxyproline, a measure of fibrosis, compared with the normal group. Grape skin and seed treatment blocked the DMN-induced increase in hepatic hydroxyproline content by 127% and 123%, respectively (Table 5), suggesting decreased collagen deposition.
Lipid peroxidation
The lipid peroxidation in the liver was measured by MDA determination in liver homogenate. As shown in Table 6, the MDA content in DMN group was higher than that of the normal group (153% of the normal) and in the DK10 and DS10 groups, it provided 70% and 90% of protection, respectively. These data indicated that both grape skins and grape seed treatment inhibited lipid peroxidation in the DMN-induced liver damage.
Discussion
Hepatic fibrosis represents a common response to chronic liver injuries, e.g., viral, metabolic, and toxic. Regardless of the type of insults, liver fibrosis is characterized by the increased production of extracellular matrix (ECM) proteins. Hepatic fibrogenesis is accompanied by hepatocellular necrosis and inflammation. HSCs are regarded as the primary target cells for inflammatory stimuli in the injured liver, and their activation may be initiated by paracrine stimuli from injured neighboring cells, including hepatocytes and Kupffer cells [16,17]. Activated HSCs have been identified as the primary source of excess ECM components in liver fibrosis [4,18].
In this study, DMN-induced increase in serum AST, ALT, ALP levels was significantly suppressed by grape skin or seeds ingestion. In chronic liver diseases, the serum albumin levels are reduced due to protein synthesis disruption in the liver. The grape skin and seed treatment blocked these DMN-induced reductions in serum and total protein. The liver occupies a central role in the metabolism of bile pigments in hepatic uptake, conjugation, and excretion phases. Excretion of bile pigments can be impaired after liver damage. The grape skin or seed treatment blocked increases in plasma bilirubin. These results suggest that grape skin or seed ingestion protects the hepatocytes from injuries and improves the liver functions of the DMN-treated rats. The damaged hepatocytes are potent sources of reactive oxygen intermediates and these compounds exert paracrine stimulation of stellate cells. Therefore, the hepatoprotective effects of grape skin and seeds may decrease paracrine stimuli, which lead to hepatic fibrosis via activated HSCs. Our present data show that DMN increased the number of α-SMA positive cells in the liver and these proliferations were suppressed by grape skin or seed ingestion and that grape skin or seeds significantly suppressed the increased tissue hydroxyproline content and collagen accumulation in the DMN-induced liver injury in rats. Taken together, these findings suggest that antifibrotic effect of grape skin and seeds may be due to, at least, suppressed HSC activation via hepatoprotective effect.
Oxidative stress plays an important role in many types of acute liver injury [19], chronic liver damage, and hepatic fibrosis [20]. Many experimental and clinical data indicate that a common link between chronic liver damage and hepatic fibrosis may be related to oxidative stress, which is associated with HSC activation [21]. Lipid peroxidation could change the properties of biological membranes, resulting in severe cell damage and play a significant role in the pathogenesis of disease. There is sufficient evidence suggesting that lipid peroxidation occur in both acute and chronic liver injuries. It has been shown that certain lipid peroxidation products induce fibrogenic cytokines and increase the synthesis of collagen by initiating HSC activation [22]. Therefore, reducing oxidative stress, which is an important stimulus to activation of HSCs, is a relatively practical avenue of intervention. Malondialdehyde is a secondary product of oxidative stress formed during lipid peroxidation. This study showed that the DMN-treated rats exhibited increased levels of hepatic MDA and that grape skin and seeds reduced this increase, respectively. These outcomes suggest that the mechanism for the hepatoprotective effects of grape skin and seeds in the development of liver fibrosis may be related to the reduction of lipid peroxidation.
Grape contains large amounts of phytochemicals, including resveratrol, quercetin, proanthocyanidins, anthocyanidin, procyanin, myricetin, kaempferol, catechin, and epigallocatechin gallate [5], which have antioxidative, anti-inflammatory, and antimicrobial activities, as well as cardioprotective, neuroprotective, and hepatoprotective effects. Most (60-70%) of grape polyphenols are found in grape seeds and the rest 30% in grape skin [6,23,24]. Among those phytochemicals, high levels of resveratrol mainly exist in the skin of grape and proanthocyanidins exist in the seed of grape. Resveratrol is reported that it has the antifibrotic effects in rat fibrosis model by CCl4 [25] and DMN [26]. Proanthocyanidins are antifibrotic in experimental biliary obstruction [27] and in the DMN-induced fibrosis model [28] in rats. The composition and amounts of the ingredients are different among the skin and seed of grape, but in this study, there is no difference in hepatoprotective and antifibrotic activities between the skin and seed treated groups. Besides to resveratrol and proanthocyanidin, lots of other bioactive components of grapes may also mediate the antifibrotic effects of grape skin and seeds, even though the contribution of these components to the antifibrotic effects is still unclear. Grape skin and seeds are usually discarded in regular dietary intake and the winery and grape juice industry. However, most of active ingredients of grape exist in skin and seed, and grape intake with its skin and seeds in the normal human diet may help maintain good health [29], especially for liver. Using these resources as active components rather than waste may have economic benefits [23].
In summary, the present study demonstrated that grape skin and seeds exhibited in vivo hepatoprotective and antifibrotic effects against liver injury induced by DMN. The mechanism appeared mostly to be mediated by inactivation of HSCs. In addition, grape skin and seeds might also produce beneficial effects by reducing oxidative stress as efficient antioxidants in DMN-treated rats, which exerted protective effects against HSC activation. Our data suggests that grape skin and seeds may be potentially useful in the prevention of the development of hepatic fibrosis.






 XML Download
XML Download