

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Cadmium (Cd) is a highly toxic environmental heavy metal and classified by International Agency for Research on Cancer of USA (IARC) as a human carcinogen (IRAC, 1993). Occurrence of toxic effects by Cd depends on the route, the concentration and the exposure time (Lopez et al., 2006). Occupational exposure to cadmium, such as working with cadmium containing pigments, plastics, glass, metal alloys and electrode material in nickel-cadmium batteries, and non-occupational exposures such as food, water and cigarette smoke induce uptake of Cd from the environment into the body through pulmonary and enteral pathways (Waisberg et al., 2003). Several population studies showed that not only occupationally exposed workers but also environmentally exposed populations can experience moderate to severe health problems due to Cd toxicity (Akesson et al., 2006; Jarup, 2002; Nordberg, 2004; Sandy et al., 2007; WHO, 2000). The detailed mechanisms for the toxic effects of cadmium are still largely unknown. However, it is thought to involve interferences with antioxidant enzymes (Hussain et al., 1987), alteration in thiol proteins (Belyaaeva & Korotkov, 2003; Li et al., 1993), inhibition of energy metabolism (Muller, 1986), alteration in DNA structure (Coogan et al., 1992) and affectation of some enzyme activities (Casalino et al., 2002; Jay et al., 1991; Watjen et al., 2001).
Chlorella vulgaris (CV) is unicellular marine algae. Chlorella contains about 55~67% protein, 1~4% chlorophyll, 9~18% dietary fiber, and an amount of minerals and vitamins (Kunimasa et al., 1999). These algae are considered to be highly resistant to heavy metals including Cd. It can also chelate heavy metal ions such as Cd (Guzman et al., 2001). These effects are thought to be mediated through metal ion-protein interactions of a variety of cellular targets. And in some cases, the metal ions-protein complex form is considered to participate in detoxification mechanisms (Goyer, 1983; Goyer, 1984; Yoshida et al., 2006). Chlorella is widely used as a food supplement in the forms of tablets, capsules, extract liquid or a food additive in Japan (Merchant & Andre, 2001). Numerous experimental studies have demonstrated various pharmacological effects of CV and CV extract, which include improvement in promotion of dioxin excretion (Yoshida et al., 2006), growth of cells, animals and plants (Han et al., 2002), and growth and development of children (Lee & Lee, 2002).
In several studies (Kim et al., 2003b; Kunimasa et al., 1999; Morita et al., 1999; Shim, 2008), when rats fed dietary chlorella simultaneously with cadmium or dioxin, chlorella intake might stimulate cadmium excretion in urine and feces, and decrease synthesis of metallothionein (MT) in liver, kidney and small intestine. However, these events mostly resulted from preventing the absorption of Cd by chlorella intake. When Chlorella vulgaris is exposed to heavy metals, metal binding MT-like proteins in the cell were induced and these proteins might be involved in the detoxification of heavy metals. (Huang et al., 2009) In this study, therefore, it was wondered that after cadmium is absorbed, to the exclusion of excretion in feces, if dietary chlorella intake could have some effects on betterment of growth and Cd detoxification. The overall goal of this study was conducted to investigate whether dietary Chlorella vulgaris (chlorella) intake would be effective on growth and Cd detoxification in rats already stressed with 8-week, high-level Cd exposure.
Materials and Methods
Experimental animals and diets
Fifty 14-week-old male Sprague Dawley rats (CD (SD) IGS, Outbred, Charles River Origin; Jung-Ang Lab. Animal, Inc., Korea) were placed in individual stainless steel wire-mesh cages in a climate-controlled room. The room had a 12:12 hr light-dark cycle, a temperature of 22-24℃ and a relative humidity of 45 ± 5%. The rats were fed a pellet diet (Samyang Co., Korea) for the first seven days (adaptation period). The rats weighed 415.03 ± 1.60 g (mean ± standard error) at the end of the adaptation period.
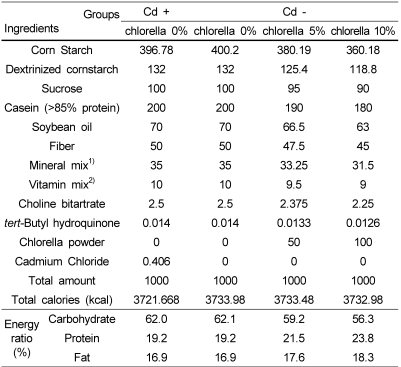
The rats were randomly divided into Cd-free group (n=10) and Cd group (n=40). They were fed the modified American Institute of Nutrition (AIN)-93G diet without or with 200 ppm cadmium (CdCl2 406 ppm, 0.04% of kg diet) for 8 weeks. For confirming cadmium-induced stress applied, all rats of the Cd free group and 10 rats of the Cd group were sacrificed. The remaining thirty rats in Cd group were then stratified according to body weight and randomly blocked into three treatment groups according to different levels of chlorella (0, 5, or 10% of kg diet, w/w) in Table 1 for the experimental period, which lasted for 4 weeks. The chlorella sample used in this study (Chlorella vulgaris) was obtained from Daesang Co. (Korea). The diets were formulated according to the nutrient content of the AIN-93G diet.
This study was conducted at the nutrition laboratory of Ewha Womans University, in compliance with the Guide for the Care and Use of Laboratory Animals. During the experimental period, the rats were allowed ad libitum access to the experimental diets and distilled water. For preventing contamination of other minerals, all instruments were treated with 0.4% ethylenediaminetetraacetic acid (EDTA) solution and washed with distilled deionized water.
Specimen collection and analytical method
Urine was collected in an acid-washed bottle treated with a few drops of toluene as preservatives using the metabolic cages during the final three days of experimental period. After collection, the bottle was filled up to 100 ml with deionized water and centrifuged at 7,000 rpm for 10 min (Supra 22K High speed centrifuge, Hanil, Korea). Aliquots of the samples were stored at -20℃ until determination of Cd concentration. Before sacrificing, animals were deprived of food for 12 hrs and sacrificed after anesthetization with diethyl ether. The liver, kidney, spleen, intestine, epididymal fat pad and perirenal fat pad were removed and weighed. The kidney and the intestine were perfused with saline and stored at -80℃ until analysis for MT determination.
Cd concentration in urine was measured by the method of Zinterhofer (Zinterhofer et al., 1971). Urine was adjusted to pH 5.5. 10% Triton X-100, 2% ADPC (Ammonium pyrrolidine-dithiocarbamate) and MIBK (4-Methyl-2-pentanone, methyl isobutyl ketone) were added to urine one after another. The solutions were mixed well, and centrifuged at 3,500 rpm for 20 min at RT (Supra 22K high speed centrifuge, Hanil, Korea). MIBK layers were treated with nitric acid and determined using AAS (Atomic Absorption Spectrophotometer, Model 6701F, SHIMADU CO., Japan). The analyses were performed at the 228.8 nm resonance line.
Determination of MT was measured by the methods of cadmium/hemoglobin affinity assay (Eaton & Toal, 1982; Eaton & Cherian, 1991). One gram of small intestine or 0.5 g of kidney was homogenized in 1 vol. of 10 mM Tris-HCl buffer (pH 7.4, 4℃) and centrifuged at 10,000 rpm for 10 min at 4℃ (Micro 17R, Micro Refrigerated Centrifuge, Hanil, Korea). The supernatant fraction was heated for 2 min in a boiling water bath. The heated samples were then centrifuged at 10,000 rpm for 2 min to remove precipitated proteins. The samples were mixed with 0.26 mM CdCl2 solution and incubated at RT for 15 min. The metal binding sites of MT were saturated with Cd during this incubation step. For removal of excessive Cd, it was precipitated by addition of rat RBC hemolysate, which was prepared following the method of Onosaka and Cherian (Onosaka & Cherian, 1982), and heat treatment in a boiling water bath for 2min. The heat treated Cd-bound hemoglobin was removed by centrifugation at 10,000 rpm for 2 min at 4℃. The amount of Cd in the heated supernatant fraction is converted from the amount of MT-bound Cd and was determined in using AAS. The analyses were performed at the 228.8 nm resonance line. The concentration of MT in each tissue was calculated by assuming that 7 g-atom of Cd is bound to each mole of thionein which has a molecular weight of 6050 by amino acid analysis.
Statistical analysis
All statistical analyses were performed by the SAS program package version 9.1. All results were expressed as mean ± standard error (SE). The data between Cd fed and Cd non-fed group were analyzed by unpaired t-test. The data between chlorella fed and chlorella non-fed group was analyzed by one-way analysis of variance (ANOVA) and differences among experimental groups were evaluated using Duncan's multiple range tests at the P<0.05 significant level. All analyses were performed in duplicate or triplicate.
Results
Food intake, body weight change and energy efficiency ratio
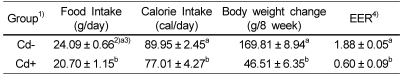
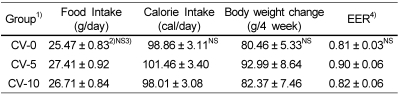
Daily food intake, calorie intake, body weight change and body weight gain/calorie intake (energy efficiency ratio, EER) were shown in Table 2 and Table 5. Daily food intake, calorie intake, body weight change and EER in rats fed Cd-containing diet (Cd+ group) were significantly lower than those in rats fed Cd free diet (Cd- group). Daily food intake, calorie intake, body weight change and EER in CV-5 group were tended to be higher than those in CV-0 and CV-10 groups, though not significantly.
Organ and tissue weights
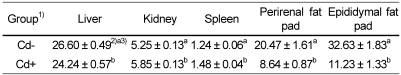
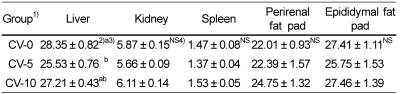
Organ (liver, kidney and spleen) and tissue (epididymal and perirenal fat pad) weights per kg body weight are shown in Table 3 and Table 6. Weight of liver, kidney and spleen in rats fed Cd-containing diet was significantly lower than those in rats fed Cd free diet. Both perirenal and epididymal fat pad weights in Cd fed group were significantly lower than those of Cd- group. Organ (liver, kidney, and spleen) and epididymal fat pad weights in CV-5 group were tended to be lower than those in CV-0 and CV-10 groups. There was no observable difference in perirenal fat pad weight between chlorella untreated group(CV-0) and chlorella fed groups (CV-5 and CV-10).
Based on the above results, the rats fed Cd containing diet were exposed to some stress. However, in rats fed chlorella containing diet, food intake, recovery of suppressed body weight gain, and organ and tissue weights were not affected compared with those fed chlorella free diet.
Cadmium concentration in urine and metallothionein (MT) concentrations in kidney and small intestine
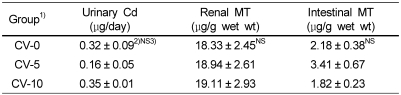
Urinary Cd concentration and MT concentrations in kidney and small intestine were shown in Table 4 and Table 7. Urinary Cd concentration in Cd fed group was significantly higher than that in Cd non-fed group. Renal and intestinal MT concentrations in the rats fed Cd containing diet were significantly higher than those in the rats fed Cd free diet, as well. Though not significantly, urinary Cd concentration in CV-5 group was tended to be lower than CV-0 and CV-10 groups. Renal MT concentration was tended to be higher in a dose-dependent manner in chlorella fed rats compared to chlorella untreated rats. Intestinal MT concentration in CV-5 group was higher than those in CV-0 and CV-10 groups.
There were no significant differences in urinary Cd and renal and intestinal MT concentrations among the groups (CV-0, CV-5 and CV-10 groups).
Discussion
Growth inhibition is the general symptom of Cd exposure (Choi & Rhee, 2001; Kim et al., 1999; Shim, 2008; Yoon & Rhee, 1994). In the present study, the rats fed Cd-containing diet had significantly lower daily food intake, body weight gain and organ and tissue weights. These results were regarded that Cd decreases food intake by reducing appetite, calorie intake along with suppressed food intake, and body weight change and EER by affecting nutritional absorption and metabolism (Itokawa et al., 1974).
Urinary Cd excretion and MT concentrations of kidney and small intestine were significantly increased with dietary Cd intake (Table 4). This result is consistent with other studies. In the previous study, compared with Cd non-fed group, urinary Cd excretion and MT concentrations in those organs were significantly increased in dietary 160 ppm Cd fed group (Shim, 2008). The other study showed that urinary Cd excretion and MT concentrations in kidney were significantly increased in rats exposed to 500 ppm Cd in drinking water for 20 weeks (Kim et al., 2003a). MT content was increased in several tissues (liver, kidney, heart and lung) in different species upon time-dependent Cd exposure (Bobillier-Chaumont et al., 2006; Bonda et al., 2004; Lange et al., 2002).
Accumulation of Cd makes such a dangerous and minimizing effect on body functions leading to kidney toxicity (Shaikh et al., 1999). Many works have been demonstrated that natural compounds or substances along with cadmium intake could facilitate cadmium excretion (Choi & Rhee, 2001; Kim et al., 1999; Kim et al., 2003a; Kim et al., 2003b; Shim, 2008; Yoon & Rhee, 1994). These studies could not be convinced that chlorella supplementation after accumulation of absorbed Cd might also be effective in Cd detoxification.
Absorbed Cd is eliminated through MT from the organism mainly via urine. The amount of Cd excreted daily in urine is, however, very small: It represents only about 0.005-0.01% of the total body burden which corresponds to a biological half-life for Cd of about 20-40 years (WHO, 2000). MT has attracted attention as a protein possibly related to metabolism and detoxification of heavy metals because it has lower affinity to the essential metal zinc but high affinity to toxic heavy metals such as Cd and Hg (Kagi & Vellee, 1960). MT was noted first for its function as a heavy metal detoxicating protein (Brouwer et al., 1993). Metal binding MT-like proteins in the Chlorella vulgaris cell were induced by exposing heavy metals and these proteins might be involved in the detoxification of those metals. (Huang et al., 2009) This showed that chlorella intake have the possibility for enhancing synthesis of metal binding MT-like proteins by capturing absorbed heavy metals in the body. Accordingly, if chlorella intake facilitates urinary Cd excretion by the above mentioned mechanism, it would be helpful in the detoxification of Cd. Thus, the aim of this study was to evaluate the effect of chlorella on cadmium detoxification after dietary Cd-induced stress in growing rats.
After stopping the feeding dietary Cd, daily food intake, calorie intake, body weight change, FER and organ weight were not affected by dietary chlorella intake. This outcome was similar to the results in several studies (Hwang, 2006; Shim, 2008) that there were no significant differences in food intake, body weight change, and organ and tissue weights among three experimental groups (chlorella non-fed group, and 5% or 10% chlorella fed group). This finding was thought that, although chlorella reported the growth stimulation in itself, cells, animals, plants, infant and child (Han et al., 2002; Kim et al., 2003b), After exposing dietary Cd, chlorella intake, regardless of its percentage, would not promote improvement of suppressed growth in rats.
Though not significant, renal MT concentration tended to rise dose-dependently. In vivo studies showed that Cd and high protein containing diet increased MT synthesis in rat organs (Kwon & Kim, 1992; Park & Kim, 1996; Revis, 1981). Since chlorella contains 55~67% protein and an amount of minerals (Morita et al., 1999), even though Cd acted as an activator of MT synthesis during the cadmium feeding period, MT de novo synthesis would be promoted during chlorella intake after given Cd stimuli, although not significant.
The effect of dietary chlorella intake on intestinal MT concentration was not a dose-dependent manner. Gropper et al. (2004) reported that divalent metal ions share their transporters and excessively ingested divalent cations such as Cu, Fe and Zn impede their absorption with each other. For high concentrations of nutrients such as minerals interfere with the absorption of each other, excessive chlorella intake, in the case of the diet containing 10% chlorella, seemed to inhibit MT synthesis.
Urinary cadmium concentration was not affected by dietary chlorella intake. Several studies presented that simultaneous intake of Cd and chlorella facilitated urinary Cd excretion and this finding resulted from inhibition of absorption but not from facilitation of excretion (Kim et al., 2003b; Shim, 2008). And a few studies presented that, when renal Cd-MT form was increased, urinary Cd excretion was facilitated (Friberg, 1984; Park & Kim, 1996). Because MT concentration in kidney was somewhat increased by chlorella intake, but not observably (Table 7), its concentration did not lead to increase urinary Cd excretion. Based on above explanation, if simultaneous intake of chlorella and Cd did not happen, Cd was already accumulated in the body and chlorella supplementation could not promote Cd excretion via urine.
On the other hand, the exact mechanism of detoxification by chlorella intake was not able to be known. Since concentrations of Cd in tissues of liver, kidney or small intestine were not measured, contribution of chlorella in MT synthesis could not be figured out. Hence, in a succeeding study, it has to additionally evaluate measurements of free Cd concentrations in blood and organs, MT-bound Cd concentrations in organs and fecal Cd excretion.
Taken together, it is thought over that, after given Cd exposure, non-simultaneous dietary chlorella intake was not likely to promote growth in animal and facilitate Cd excretion via urine through detoxification of Cd-MT like proteins.

 XML Download
XML Download