

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The digestion-resistant fractions (RFs) consisted of resistant starch (RS) and non-starch polysaccharides (NSP) have important implications for diabetes, colon cancer and blood lipid chemistry (Anderson et al., 2000; Hinggins et al., 2004; Leeuw et al., 2004; Leu et al., 2002). The potential mechanisms for this health include the promotion of beneficial microflora that uses NSP of RF as a substrate in colon. And the health benefits like RS is a reduced glycemic index. Particularly, RF was found to have great effects on various disease risks with resistant starch 1 (RS1), which is physically trapped in milled grains and inaccessible to digestive enzyme after 18 hrs incubation (Annison & Topping, 1994; He et al., 2005). Also, the RF feeding had the increased the bile acid excretion in mice stool samples of apolipoprotein deficient mice after feeding RFs (Han et al., 2006).
Several studies have shown that ungelatinized starch, resistant starch 2 (RS2) of RF, which is abundant in raw potato, banana, and high amylose maize starch, are not degraded by digestive enzymes after 4-5 hrs incubation in vitro (Mccleary & Monaghan, 2002). Heating of plant food alters the physical and physiological properties of RF, which, in turn, affects the quality and quantity of RS and NSP. RF from unheated source, so-called Saeng-shik in Korea, has more RS and shows more effectiveness in lowering diabetes and cardiovascular disease risk compared to RF from processed sources (Park et al., 2003). Soybeans widely ground and consumed in various regions of the world, are rich and relatively inexpensive sources of proteins and carbohydrates. Its carbohydrates are composed of starch and nonstarch polysaccharides as resistant starch, cellulose, hemicellulose, and pentosan. It was known that they affected colon cancer, obesity and cardiovascular disease (Han et al, 2004; Lee et al., 2006). However, the change in the amount of RS and NSP of soybean during heating may be partly attributed to the redistribution of RF. Also, the governing factors for the physiological functionality of RF have not been determined.
In the present study, the effects of change of functional factors (RS1 and NSP) of RF from soybean on bile acid transport were investigated.
Material and Methods
Preparation of RFs
Soybeans [Glycine max(L.) Merr.] was purchased from Sunhan Nonghyup in Korea. Fat of soybean were defatted before preparation of digestion-resistant fractions (RFs) by hexane. RFs were prepared using a modification of Goni et al. (1996). Soybeans were finely ground before heat treatment and autoclaved under standard conditions (121℃, 15 min, 1 atm). Heat-treated and raw, unheated samples were then processed further. First, proteins in 10 g of samples were digested by incubation with pepsin in 1000 mL of KCl-HCl buffer (pH 1.5) for 60 min at 40℃. After protein digestion, carbohydrates were digested by adding 900 mL of α-amylase solution (4 mg/100 mL) in tris-maleate buffer (25.5 g/ 550 mL, pH 6.9) for 18 hrs at 40℃. After two digestion reactions, the samples were centrifuged (3000 × g, 15 min), the supernatant was discarded, and the pellet was washed three times with distilled water. The pellet was then incubated in a series of alkaline and acidic solutions for 30 min at room temperature (KOH [67.3 g/ 300 mL], followed by HCl [14.0 mL/ 550 mL]). The solution was further incubated with sodium acetate buffer (9.8 g/ 300 mL, pH 4.75). Finally, the residuals were digested with amyloglucosidase (Sigma A-7255) for 1 hr at 60℃. Each digestion reactions were performed with appropriate shaking. The final sample was washed, centrifuged (3000 × g, 15 min), and the lyophilized pellet was used as an RF in the experiments. Two types of RFs were prepared: a raw, unheated RF (RRF) and a heated RF (HRF) from soybean (SOY). The RFs were designated as RRF-SOY, and HRF-SOY, respectively.
Retarding effect of digestion-resistant fraction in vitro on bile acid transport
Retarding effects of digestion-resistant fractions (RFs) on in vitro bile acid transport were investigated by dialysis (Sigma D7884: M.W. cut-off<1,200) using a glucose transport model. 0.2 g of RF in 6 mL of phosphate buffer (pH 7.0) with sodium azide (0.65 g) and taurocholic acid (0.05 g) were added to a dialysis tube. The dialysis bag was then placed in a 150 mL capped cylindrical container containing 100 mL of sodium azide and shaken at 75 rpm in a 37℃ water bath for 24 hrs. Aliquots of 1 mL of the dialysate were removed at regular intervals, and assayed for taurocholic acid content according to others (Ju et al., 2003; Lee et al., 1996; Lee et al., 2003).
X-ray diffraction
X-ray diffraction analysis for digestion-resistant fractions (RFs) was performed with X-ray diffraction image processor (Bruker D8 discover, Germany). Analysis parameters were 40 kV and 40 mA; Cuk radiation λ = 0.154 nm. The samples were scanned though 2θ range from 5° to 40°.
Content of cellulose and hemicellulose
Cellulose was analyzed according to Kim et al. (1986). Digestion-resistant fractions (RFs) were hydrolyzed by a cellulase mixture [Trichoderma viride cellulase: Aspergillus niger cellulase, 10:1 (W/W)]. One hundred twenty mg of the cellulase mixture were added to 500 mg of RFs and 15 mL of sodium acetate buffer (0.49 g/15 mL, pH 4.8). The mixtures were allowed to react in a 50℃ shaking water bath (120 rpm) for 1 hr. Then the mixtures were centrifuged (3,500 rpm, 20 min) and analyzed for reducing sugars.
Cellulose content of RFs was determined by calculation of reducing sugar content using a DNS method (Lee et al., 2005). In brief, 0.5 mL of supernatant and 1.5 mL of DNS reagent (1,000 mL distilled water + 7.5 g dinitrosalicylic acid + 14 g NaOH + 216 g potassium sodium tartrate + 5.4 mL phenol + 5.9 g Na2S2O5) were mixed well. After mixing, the samples were allowed to stand for 5 min in boiling water, and then cooled to room temperature in ice water. The absorbance of the samples was measured at 550 nm and the cellulose of samples was calculated according to the following equation:
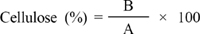
A: the absorbance of reducing sugar produced from reagent-grade cellulose.
B: the absorbance of reducing sugar produced from samples.
Hemicelluloses in RFs were measured according to others (Lee et al., 2004; Nagata et al., 2001). First, to remove the amount of protein in RFs from soybean, One hundred mL of sodium chloride (5 g/ 95 mL) were added to 1 g of sample and homogenized for 20 min. The samples were then centrifuged at 10,000 rpm for 10 min. 100 mL of alcohol (80 mL/ 100 mL) were added in the precipitate, extracted, and filtrated. Alcohol-insoluble RFs were extracted by KOH (22.4 g/ 100 ml) and centrifuged. The pH of the supernatant was then adjusted to pH 4 with hydrochloric acid. The residue of hemicelluloses was lyophilized and the yield was calculated.
Pentosans
The total pentosans were analyzed according to others (Hashimoto et al., 1987; Delcour et al., 1999) with some modification. The samples were weighed (10 mg) and 2 mL of HCl (14.0 mL/ 550 ml) were added. The mixtures were then hydrolyzed at 100℃ for 3 hr. After cooling, the samples were neutralized by adding 2 mL of sodium carbonate (0.21 g/ml). Saccharomyces cerevisiae yeast (25 mg/mL of sodium phosphate buffer [0.16 g/ mL], pH 7.0) was added and the samples were incubated in a shaking water bath at 37℃ for 1 hr. The mixture was then centrifuged at 3000 × g for 10 min. The supernatant was analyzed by the Orcinol-HCl method (Albaum & Umbreit, 1947).
Analysis of monosaccharides in digestion-resistant fractions (RFs)
Monosaccharide composition of digestion-resistant fractions (RFs) from soybean was determined by using of a high performance anion exchange chromatography (HPAEC) (Bio-LC; Dionex Co., Sunnyvale, California, U.S.A). Ten mg of RFs were hydrolyzed in 1 mL of trifluoroacetic acid (TFA, [57.2 mg/mL]) for 2.5 hr at 100℃. An analytical column for carbohydrate detection (CarboPac PA1; Dionex Co., Sunnyvale, California, USA) and an electrochemical detector (ED50A; Dionex Co., Sunnyvale, California, USA) were used. Separation of the various monosaccharides was achieved using eluent A (NaOH [0.72 g / mL]) followed by application of a linear gradient for 30 min using eluent B (NaOH [8 g/ mL]) (Seo et al., 2004).
Statistical analysis
The results are expressed as mean ± standard division (n=3). An analysis of variance (ANOVA) was performed, and the differences among the samples were determined by Duncan's Multiple Range Test using the Statistical Analysis System (Version 9.13, SAS Institute, GA, USA). P values<0.05 were considered significant.
Results
Chemical composition of digestion-resistant fractions (RFs)
Raw, unheated (R) and heat-treated (H) digestion-resistant fractions (RFs) were prepared from soybean (SOY). The yield of RF was approximately 5 g per 100 g unheated materials. However, the yield was markedly reduced after the heat treatment (2.4 g per 100 g of soybean). The composition of two types of RFs was shown in Table 1. Carbohydrate, protein, and lipid contents of RFs were nearly zero. The RF from raw, unheated soybean had relatively high level of resistant starch (RS) compared to that of heated sample. After heating, the content of RS reduced, but the fiber was vice versa.
Retarding effect of digestion-resistant fraction in vitro on bile acid
The digestion-resistant fractions (RFs) from soybean were retarded bile acid transport (Fig. 1). In only bile acid solution (control) was dialyzed and its transport continually increased up to 18 hrs. But samples of RFs including were significantly retarded the bile acid transport. Also, digestion-resistant fraction from raw, unheated soybean (RRF) induced low level of bile acid transport than that of digestion-resistant fraction from heated soybean (HRF) up to the limit time, although level of transport increased rapidly in early stage of dialysis (<4 hrs). The decrease in transport level brought by adding RRF-SOY and HRF-SOY were 26.7 and 48.2% for bile acid. This retarding effect is in accordance with results of previous in vivo experiments (Lee et al., 2003). As shown the Table 1, heat treatment changed compositions of RF. So it seems to be that the composition of RFs from soybean affected the functionality of RF.
X-ray diffraction patterns
The X-ray diffraction patterns of digestion-resistant fractions (RFs) were showed in Fig. 2. The RFs from soybean were characterized by peak at different angles (2θ) corresponding to resistant starch (RS) content. The structures of RFs were influenced by heating. The peak of raw soybean starch as a control of X-ray diffraction had the strong intensity at diffraction angles of 12.0 and 20.0°. The RF from unheated soybean (RRF-SOY) showed similar the peak intensity to raw soybean starch, control. However, the RF from heated soybean (HRF-SOY) was characterized by a peak at a diffraction angle (2θ) of 19.8°. The X-ray pattern of HRF-SOY were reduced peaks at diffraction angles of 12 and 20° compare to RRF-SOY.
Cellulose and hemicellulose
Cellulose and Hemicellulose content of digestion-resistant fractions (RFs) from unheated and heated soybean was shown in Fig. 3. The RFs of two samples had cellulose over 20%. The RF from unheated soybean (RRF-SOY) had significantly high level of cellulose content than RF from heated soybean (HRF-SOY). However, in case of hemicellulose, the hemicellulose content of RRF-SOY and HRF-SOY showed different trend compared to the results of cellulose of RFs. Hemicellulose of RFs from soybean had over 30%. Hemicellulose of RRF-SOY was low level than HRF-SOY.
Pentosans of digestion-resistant fractions from soybean
Pentosan content of digestion-resistant fraction (RF) from unheated and heated soybean was shown in Fig. 4. Pentosan content of RF from unheated soybean (RRF-SOY; 29.6%) was significantly higher value than RF from heated soybean (HRF-SOY; 25.1%). This was similar trend like those of RS and cellulose in RRF-SOY. Pentosan is a polymer of various pentoses, arabinose, xylose, and ribose and that sort of things. Particularly, arabinose and xylose were well known to have the physiological functional components (Marlett & Fischer, 2003). So, this result could be explains a correlation with the retarding effect of RF on bile acid in vitro.
Monosaccharides in digestion-resistant fractions
Peaks for rhamnose, arabinose, galactose, glucose, xylose, mannose, and fructose in digestion-resistant fraction (RF) from unheated soybean (RRF-SOY) and RF from heated soybean (HRF-SOY) were assigned by comparison with standard (Fig. 5 and Table 2). In RRF-SOY, glucose, galactose, and arabinose were found to have the highest levels at 32.5, 25.2, and 15.0%, respectively. Whereas in HRF-SOY sample had glucose, galactose, and arabinose levels at 1.9, 30.1, and 31.3%. On the other hand, arabinose is a component of hemicellulose. But the hemicellulose of HRF-SOY was higher than did it RRF-SOY. These results indicate that the functional factors of RF from soybean on retarding effect on bile acid might be not much influenced by arabinose. And the retarding effect of RF on bile acid transport might be big influenced by glucose as a component part of RS. Also, xylose content of RRF-SOY was higher 35.2% than that of HRF-SOY.
Discussion
RFs are known to have an affect on various diseases. Particularly, effect of RFs from soybean on cholesterol metabolism was reported (Han et al., 2004). In this study, we examined the retarding effect of heat treatment of digestion-resistant fraction from soybean on retarding of bile acid transport in vitro. In consequence, heat treatment had affected to RFs content and it's functionality on retarding bile acid. After heating, RS in RFs content were markedly reduced. And the levels of cellulose and pentosan in RFs from unheated soybean were higher, whereas the levels of RS in RFs from heated soybean were reduced. The retarding effect showed difference with different RFs content. We conformed reduction of RS in RFs after heating by X-ray diffraction. The peak of RS was reduced in RF from soybean so that NSP peak was distinguished after heating, relatively. These results seem to be that the RS in RFs was melted by heating. The X-ray diffraction depended on the chain lengths making up the amylopectin lattice, amylose and the density of packing within the granules. Therefore, X-ray diffraction had affect heat treatment (Sajilata et al., 2006; Xie et al., 2006). And Sievert and et al reported that effect of autoclaving and autoclaving-cooling cycles of RS. After autoclaving, the peak of X-ray diffraction was lower than those native starches and control. Also, autoclave cycle was increased as the peak of X-ray diffraction was decreased (Siever et al., 1991). Therefore HRF-SOY may have also to be more affected by NSP than RRF-SOY because RS was melted by heat treatment (Ruan et al., 2004). These were supported by result of cellulose, hemicallulose and pentosan. In case of hemicellulose, that of RRF-SOY was lower level than HRF-SOY. Whereas cellulose content of RRF-SOY was higher level than HRF-SOY. So the RRF-SOY of low hemicellulose content or high cellulose content will affect on the physiological function (Takahashi et al). The chemical structure of cellulose is different. Cellulose is linear polymer of β-1,4-linked glucose units. Hydrogen bonding between sugar residues in adjacent chains imparts a crystalline microfibril structure. Hemicelluloses are a cell wall polysaccharides containing backbones of β-1,4-linked pyranosides sugars, but differ from cellulose in that they are smaller in size, contain a variety of sugars, and are usually branched. Moreover, the researcher reported that these structure characteristic of hemicellulose from soybean influenced on serum antibody levels and activation of microphage in rats (Nagata et al., 2001). And cellulose seems to play a role in bile acid-binding when it stabilizes the cell wall architecture after the digestion process (Dongowski 2007). Also, different sugar content in RFs supported that effect of thermal processing of soybean on the sugar composition of RFs (Periago et al., 1997). In sugar composition, the glucose and xylose content of RF from unheated soybean was higher than the RF from heated soybean. The xylooligosaccarides is well known to have functional monosaccharide on retarding effect of bile acid. Moreover, the retarding effect of RF on bile acid also had been affected by the difference of sugar content (Vazquez et al., 2000).
The level of RS, pentosan, glucose, and xylose were inversely correlated to that of the in vitro retarding effect on bile acid. Therefore, this study suggests that heating soybean has affected to the RF of functional components. Also, we think that the RS and the monosaccharide composition like xylose are important factors for retarding effect on bile acid transport.






 XML Download
XML Download