

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
According to American Heart Association Report, cardiovascular disease (CVD) is a major cause of death in US and the death rate of 35.2% is accounted from CVD in 2005 (Rosamond et al., 2008). Elevated plasma cholesterol is one of the major risk factors in developing cardiovascular diseases, and an intervention study showed that reduction of total cholesterol by 11.5% reduced CVD events by 33% (Nakamura et al., 2007). Statin, a HMG CoA reductase inhibitor is recognized as one of the most popular drug lowering cholesterol in human. Among US adults aged ≥ 40years with hypertension, 14.5% are taking statin medication in NHANES (1999-2002) study (King et al., 2007). Statin is known to deplete coenzyme Q10 (CoQ10) that shares cholesterol biosynthetic pathway, resulting in decreased antioxidant capacity. The reduced form of CoQ10 (CoQH2), ubiquinol in plasma membrane functions as radical scavenger, protecting cell membrane from oxidative damage (Mabuchi et al., 2005). Nitric oxide (NO) that plays a dual role as deleterious radical species and beneficial species can be induced by statin (Fontaine et al., 2002). Statin dependant endothelial nitric oxide synthetase (NOS) affected Na-K ATPase and Na-K cotransport in rat renal tubule (Varela et al., 2004). Statin may influence membrane ion channel by changing membrane lipids, thereby affecting membrane fluidity. Membrane fluidity of erythrocyte was increased with decreasing plasma cholesterol by statin therapy (Broncel et al., 2007). NO inhibited platelet aggregation through cyclic GTP pathway in simvastatin treated rabbit (Chou et al., 2008). A NO donor, nitro-L-arginine methyl ester (L-NAME) or antioxidant quercetin and catechin exerted antiplatelet effects by inhibiting NADPH oxidase and increasing NO (Pignatelli et al., 2006). Betalain pigments which are present in red beet (Beta vulgaris L.) root and prickly pear cactus (Opuntia ficus indica) fruit are recently recognized as bioavailable natural radical scavengers. Tesoriere et al. (2005) reported that betalain metabolite, betanin and indicaxanthin were observed in LDL fraction at 3 hours after ingestion of prickly pear cactus fruit and the resistance of LDL particles to ex vivo induced oxidation was increased. Cationized antioxidants in red beet inhibit lipid peroxidation in membranes or linoleate emulsion and prevent H(2)O(2)-activated LDL oxidation at relatively low concentration (Kanner et al., 2001). Quercetin which is abundant in most fruits and vegetables is the major type of flavonoid in onion (Allium cepa). Yamamoto et al. (2005) have shown that the green-leafy Welsh onion reduced superoxide generation by suppressing the angiotensin II production and then the NADH/NADPH oxidase activity, increasing the NO availability in the aorta and consequently lowering blood pressure in the rats fed with a high fat diet. Continuous ingestion of onion soup inhibits collagen induced platelet aggregation in human by inhibiting collagen stimulated tyrosine phosphorylation (Hubbard et al., 2006)
We hypothesized that statin therapy may affect some cardiovascular related parameters such as erythrocyte efflux channels, platelet aggregation besides cholesterol lowering effect by way of its own function on erythrocyte Na channel and platelet aggregation or by depletion of CoQ10 and induction of NO, and antioxidant rich foods such as onion and beet may reverse or interact statin action on these parameters.
Materials and Methods
Animals and diets
Forty of twelve weeks old Sprague Dawley rats (Orient Bio Co., Ltd, Korea) were divided into four groups and fed the following diets: Control; 0.5% cholesterol based diet: Simvastatin (Choongwae Pharm. Co., Korea); the control diet plus 2 mg/kg BW simvastatin: Sim-onion; the simvastatin diet plus 5% onion powder: Sim-beet; the simvastatin diet plus 5% beet powder. Rats had free access to water and were housed individual cages in a room maintained at 20-25℃ with a 12 hour dark-light cycle. After 4 weeks ad libitum feeding, blood samples were obtained by cardiac puncture into heparinized vacuum tubes, and platelet aggregation and erythrocyte Na efflux were performed with fresh blood. Liver samples were prepared for microscopic examination and plasma and liver samples were stored at -70℃ for later assays.
Platelet aggregation
Platelet aggregation was measured using a Chronolog Whole Blood Aggregometor (model 500-Ca, Havertown, Pennsylvania, USA), of which the instrumental principle is based on the increase in impedance (Ω) across two platinum electrodes as platelet aggregation proceeds. The whole blood was diluted with isotonic saline (1:4) to give platelet concentration of approximately 200,000 platelets/µl. Adenosine diphosphate (ADP, 2 µM) was added to initiate aggregation, and three readings of impedance changes were averaged for each rat.
Plasma and liver lipid assays
Plasma total cholesterol, HDL-cholesterol and triglyceride were assayed using enzymatic kits (Asan Pharmaceuticals, Korea). Ten µl of plasma was used for the assays of total cholesterol and triglyceride. A 200 µl sample of plasma was incubated with dextran sulfate to precipitate apo B containing lipoprotein and 50 µl of the supernatant was used for HDL cholesterol.
Liver lipids were extracted by a modified Folch method (Folch et al., 1957). One gram of liver tissue was homogenized for 5 min in 6ml of Folch solution [chloroform (2): methanol (1)] and 2ml H2O. After centrifugation for 10 min, the lower phase that contains liver lipids was separated. Lower phase of lipid fractions was assayed after treating with triton X-100:chloroform (25 µl:475 µl) for total cholesterol or with methanol for triglyceride, using enzymatic kits (Asan Pharmaceuticals, Korea).
Na Efflux Channels
Red cell preparation:
Blood was centrifuged at 1,000 ×g for 10 minutes, and the plasma and buffy coat were removed. Red blood cells were washed 5 times with a cold isotonic washing solution [150 mM choline chloride, 10 mM Tris-4 morpholinopropane sulfonic acid (MOPS), pH 7.4 at 4℃], and centrifuged at 1,000 ×g for 5 minutes after each wash. The RBC pellet was resuspended in the choline chloride washing to give 40-50% hematocrit, which was also measured. A 50ul aliquot of the RBC suspension was added to 5ml of 0.025% acationox (a metal free detergent, Scientific products, McGraw Park, Illinois, USA) to be used for determination of intracellular Na concentrations.
Na efflux:
Four ml each of the RBC suspension were added to 40ml MgCl2 medium with and without ouabain (70 mM MgCl2, 10 mM KCl, 85 mM sucrose, 10 mM glucose, 10 mM Tris MOPS pH 7.4 at 37℃, 1 mM ouabain) for determination of Na efflux via Na-K ATPase. Two ml of the RBC suspension was added to 40 ml of choline chloride medium with and without furosemide (150 mM choline chloride, 10 mM glucose, 1 mM ouabain, 10m Tris-MOPS pH 7.4 at 37℃, 1 mM furosemide) for determination of Na efflux via Na-K cotransport. The RBCs in each medium were mixed and aliquoted into 12 tubes. Tubes in duplicate were transferred to an ice bath after incubation at 37℃ in a shaking water bath for 0, 2, 4, 6, 8, and 10 minutes for Na-K ATPase and 0, 10, 20, 30, 40 and 50 minutes for the Na-K cotransport. Tubes were centrifuged at 1,000 ×g for 5 minutes and then the supernatant was removed and measured for Na concentration using Atomic Absorption Spectrophotometer (Shimadzu model AA6701F).
Calculation:
Ouabain sensitive Na-K ATPase can be blocked by ouabain and furosemide sensitive Na-K cotransport can be blocked by furosemide. Sodium efflux via Na-K ATPase was calculated as the difference between the efflux into MgCl2 medium with ouabain and the efflux into MgCl2 medium without ouabain; Na-K cotransport was calculated as the difference between the efflux into choline chloride medium with furosemide and the efflux into choline chloride medium without furosemide; Na passive leak was calculated as the efflux into choline chloride medium that contained ouabain and furosemide. For the determination of the efflux via Na-K ATPase with 4 ml erythrocyte suspension added to 40 ml MgCl2 medium, the following calculation was used (Kang et al., 1990).
Na µg/(ml×min)×60 min×µmole/23 µg×[44 ml-(4 ml×hct)/(4 ml×hct)]×=µmole/ml rbc/hour (mmole/ℓ rbc/hour)
For the determination of intracellular Na concentration with 50 µl red cell suspension added to 5ml 0.02% acationox, the following calculation was used;
Na µg/(ml×min)×60 min×µmol/23 µg×101/hct = µmole/ml rbc (mmole/ℓ rbc)
Histological examination
Two parts of liver and aorta samples each were obtained from four rats per group. After being washed with saline, samples were fixed in 10% formalin for 48 hours and dehydrated in xylene for 18 hours. Samples were then embedded in paraffin and cut into 4-5 um cross section. Tissues were stained with hematoxylineosin for microscopic observation.
Results
Weight gain and food efficiency
The final body weight along with average daily gain (ADG) was not different among groups (Table 2). The feed efficiency ratio (FER) tended to be a little higher in fiber containing SIM-onion and SIM-beet groups. There was no difference in liver/body weight ratio (LW/BW).
Plasma and liver cholesterol and triglyceride
All groups fed diets containing simvastatin were significantly increased in plasma total cholesterol and the difference between the control and SIM group was significant (p<0.01) (Table 3). Plasma total cholesterol in SIM-onion and SIM-beet groups was significantly decreased compared with SIM group (p<0.05). HDL-cholesterol was significantly increased in SIM-beet group compared with other groups (p<0.05). There was virtually no difference in plasma triglyceride among groups.
Liver total cholesterol was not different between any two groups, but liver triglyceride was significantly different between SIM group and SIM-onion group (p<0.05).
Whole blood platelet aggregation
Hematocrit was not different between any two groups (Table 4). Platelet aggregation in both the maximum aggregation and initial slope was significantly increased in SIM-onion group compared with simvastatin group (p<0.05).
Na efflux channels
Erythrocyte Na efflux through Na-K ATPase was significantly decreased in SIM group compared with the control, SIM-onion and SIM-beet group (p<0.05). Na efflux through Na-K cotransport was also decreased in SIM group compared with the control and SIM-onion group. Na passive leak was significantly increased in all groups treated with simvastatin compared to the control (p<0.05). The total Na efflux was decreased significantly in SIM group compared with all other groups (p<0.05). There was no statistical difference in intracellular Na among groups.
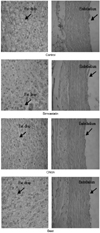
Histological examination of liver and aorta
Despite of higher plasma cholesterol, fat accumulation of liver tissue in simvastatin and SIM-onion groups appeared less than that of the control (Fig. 1). Endothelium of aorta tissue of the control looked more rumpled than that of groups treated with simvastatin.
Discussion
Higher food efficiency ratio in SIM-onion and SIM-beet can be attributed to short chain fatty acids produced from fermentation of fibers in onion and beet. Some of undigested natural fiber can be fermented to propionate and acetate and absorbed in the large intestine (Lambo-Fodje et al., 2006). Plasma total cholesterol was increased in all groups treated with simvastatin opposing to our expectation. Condo et al. (1996) reported atorvastatin at 1, 3, 10 or 20 mg/kg BW in guinea pig decreased plasma and liver cholesterol with reduced HMG CoA reductase in a dose-dependent manner. Pravastatin or fluvastatin at dose of 2 mg/kg or 4 mg/kg BW respectively did not change plasma total cholesterol in rats (Chen et al., 2003). Pravastatin at high dose as 40 mg/kg or fluvastatin 20 mg/kg BW (Bełtowski et al., 2002) in Wistar rats and rosuvastatin at 20 mg/kg Bw in SHR rats (Susic et al., 2003) had no effect on plasma HDL, total cholesterol and triglyceride. However, simvastatin 2 mg/kg BW we used in the present study did act on cholesterol metabolism causing an increase in plasma total cholesterol and a decrease in HDL cholesterol. Simvastatin, 80 mg/day or 1.8 mg/kg BW per day in human is the maximal approved dose and effective in lowering cholesterol (Lutjohann et al., 2004). Rat model may differ from human and guinea pigs in the cholesterol synthesis compartment, the plasma distribution of cholesterol and the processing of lipoprotein in plasma compartment. Unlike human in whom approximately 65% of total endogenous cholesterol is synthesized in extrahepatic tissues, the major site of cholesterol synthesis in the rat is liver (West et al., 2004). According to Fernandez (2001), plasma LCAT (lecithin-cholesterol acyltransferase) and CETP (cholesteryl ester transfer protein) in human and guinea pig are very active, while those enzymes in rat or mice are negligible therefore the plasma cholesteryl ester in rat is produced by liver ACAT (Acyl CoA-cholesterol acyltransferase) and released. The majority of circulating cholesterol in human and guinea pig is transported in LDL, but cholesterol in rats is carried by HDL fraction (Fernandez, 2001). West et al., (2004) reported that simvastatin is a lipophilic drug, which crosses membrane layers and influences cholesterol synthesis in extrahepatic tissues as well as in the liver, but there was a differential effect among species on HMG CoA reductase activity and mRNA synthesis. The different action mode among species can explain the discrepancy in hypocholesterolemic effect of statins. Onion and beet were effective in reducing plasma total cholesterol, which may be attributed to the fiber in onion and beet. Short chain fatty acids (SCFA) produced by microflora in the large intestine suppress cholesterol synthesis in rat liver and intestine (Hara et al., 1999), and SCFA produced from sugar beet fiber decreased plasma cholesterol by interrupting enterohepatic bile circulation (Hara et al., 1998). S-methyl cysteine sulfoxide in onion family suppresses HMG CoA reductase (Kumari & Augusti, 2007). Onion and beet in the present study showed a favorable effect of increasing HDL-cholesterol which is beyond any reasonable explanation. There is a report that daily consumption of 300 gram tomato for one month increased HDL cholesterol in human (Blum et al., 2006).
Induction of nitric oxide (NO) by statin is one factor for suppressing platelet aggregation (Chou et al., 2008), while depletion of plasma antioxidant ubiquinol by statin therapy is another factor for activating platelet aggregation (Pignatelli et al., 2006). Theoretically, statin itself could hardly affect platelet aggregation as shown in the present study. Platelet aggregation may be controlled by much more complicated mechanism than we thought. Most of previous studies have reported that onion has antiplatelet effect (Briggs et al., 2001; Hubbard et al., 2006), but onion in the present study increased platelet aggregation in the initial slope and the maximum. Thiosulfinates have been implicated as a principle source of the antiplatelet property of raw onion and garlic juice (Briggs et al., 2001). Hubbard et al. (2006) showed that antioxidant quercetin in onion is involved in tyrosine phosphorylation in the signaling pathway causing reduced platelet aggregation. Antiplatelet compounds such as herb extracts can suppress platelet activity in vitro at high concentration but at low concentration, those can stimulate platelet aggregation. Healthy platelets can be dissociated from their aggregate under certain physiological condition. Platelet dissociation and survival after aggregation depends on the concentration of platelet suppressing compounds in medium (Chew et al., 2001). Previous studies (Briggs et al., 2001; Chew et al., 2001) that had antiplatelet effects of onion were conducted in the in vitro system, while the effect of onion on platelet aggregation in the present study was performed in vivo. Differences in experimental protocols and species must be taken into consideration for interpretation on platelet activity.
Cholesterol lowering agent statin may decrease the membrane ratio of cholesterol/phospholipid, which may cause increase in membrane fluidity and increase in Na-K ATPase. Higher membrane cholesterol and lower Na-K ATPase were observed in the erythrocyte of hyperlipidemic patients (Broncel et al., 2007). Broncel et al. (2007) reported that fluvastatin at 80 mg/day in human decreased plasma total cholesterol and triglyceride and increased erythrocyte Na-K ATPase and Na-K cotransport. Statin in the present study rather increased plasma cholesterol, consequently increased membrane rigidity, and decreased Na-K ATPase. Statin induced nitric oxide synthetase (NOS) increasing cellular NO which in turn activates Na-K ATPase (Varela et al., 2004). On the other hand, Bełtowski et al. (2007) reported that NO decreased renal Na-K ATPase by stimulating cGMP and superoxide radicals and increased Na-K ATPase by depleting NO in leptin induced obese rats. Exposure of enterocytes to a NO donor, S-nitroso-N-acetlypenicillamine (SNAP), induced enterocyte injury in cultured intestinal epithelial cells, causing decrease in Na-K ATPase (Suzuki et al., 2005). This discrepancy in the effect of NO and oxygen radical on Na-K ATPase may be attributed to the dual effects of NO. Quercetin rich in onion and betalain rich in beet are recognized to be bioavailable antioxidants and prevent membranes from damage by free radicals. Oxidative damage of membrane causes alterations on membrane channels. Decrease in platelet membrane fluidity and Na-K ATPase were observed in patients with ischemic stroke (Nanetti et al., 2008).
In conclusion, despite reports that statins at even high dose as 20-40 mg/kg did not affect plasma total cholesterol in rats or mice, simvastatin at 2 mg/kg BW in the present experiment was enough to act on cholesterol metabolism, increasing plasma total cholesterol in rats. This dose is comparable to the maximum approved dose effectively lowering plasma cholesterol in human. The discrepancy on these results can be explained by the species difference in the metabolic pathway of cholesterol metabolism. Unlike antiplatelet action of onion in previous studies (Briggs et al., 2001; Chew et al., 2001; Hubbard et al., 2006), onion in the present study acted not as platelet suppressant but as stimulant where the platelets with high dissociation would have high survival rate after aggregation. In this study, simvastatin, which is a structural analog of HMG CoA thereby inhibiting HMG CoA reductase, increased plasma cholesterol and reduced Na-K ATPase and Na-K cotransport, and consequently decreased the total Na efflux. The results indicate that simvastatin apparently plays different roles in the cholesterol metabolism in rat differently from those in human.





 XML Download
XML Download