

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Hypertension is the single largest cause of death and is responsible for 9.4 million of deaths in worldwide [1]. It is defined as systolic blood pressure (SBP) over 140 mmHg and/or a diastolic blood pressure (DBP) over 90 mmHg [12] . Blood pressure is regulated by the renin-angiotensin-aldosterone system (RAAS), which is involved in regulating blood volume and systemic vascular resistance. RAAS is activated by low sodium concentration and glomerular filtration pressure. RAAS plays a role in reabsorption of water, blood vessel contraction, and Na+ reabsorption via angiotensin II and aldosterone [3]. Dietary salt intake is an important factor in the development of hypertension [4]. When the plasma sodium concentration increases, intracellular fluid is osmotically transferred to the extracellular space, stimulating the thirst and resulting in an increase of extracellular fluid volume. Generally, when thirst is stimulated, water consumption increases, resulting in increased total body fluid and blood pressure [56]. Consequently, the WHO recommends a salt intake less than 5 g per day. Therefore, the Korea Food & Drug Administration (KDA) not only reduced the daily recommended salt intake, but also the daily allowance of Korean fermented food containing a large amount of salt (kimchi 1 serving size: 40 g (Na+: 312.08 mg)). Ganjang (traditional Korean soy sauce) is widely used in sauces as a seasoning in Korea, and salt-free or low-salt ganjang was reported to have anti-colitis [7], anti-cancer [8], and anti-hypertension [910] properties. A recent ganjang study used the peptide from the stem of salt-free/low-salt ganjang. To our knowledge, a study using high-salt ganjang has not been reported.
Therefore, the biological effects of ganjang as the regulation system of blood pressure were investigated in an animal model.
MATERIALS AND METHODS
Materials
Ganjang was manufactured by the Sunchang Sauce Corporation (Sunchang-gun, Jeollabuk-do, Korea) as fig. 1. Ganjang was matured for 6 months. The salinity of ganjang and salt water (NaCl; Samchun, Pyeongtaek-si, Gyeonggi-do, Korea) was adjusted to 8% [1112] by Mohr's method [13]. The amount of sodium and potassium ingested by the animal was shown in Table 2. The control group orally administrated the same amount of distilled water (DW) as control.
Animal study
Five weeks male Sprague-Dawley rats (Central laboratory animal Co., Seoul, Korea) were housed in semi-SPF facilities with temperature (25 ± 2℃) and humidity controlled under 12-hr light/dark cycle conditions. Rats were randomly divided into 3 groups according to similar body weight and SBP; Control (CT), NaCl (NC), and Ganjang (GJ) after 1 week of adaptation (n = 8). The diet was given to rodent diet (Research Diets, Inc. New Brunswick, NJ, USA) and animals were orally administrated (10 mL/kg body weigh/day) for 9 weeks. SBP was measured every week in conscious rats using tail-cuff method (BP-2000, Visitech Systems, Inc., Apex, NC, USA) after 6 hours of oral administration. In last 4 weeks, rats were housed in metabolic cages for 24 hours per week, and dietary intake and water intake were measured. Furthermore, urine and feces were collected for analysis. The rats were anesthetized at the end of 9 wk experimental period. Blood was collected and centrifuged at 4,000 rpm for 15 min at 4℃, and organ tissues were harvested. The serum and all tissues were stored at -70℃ until use. All animal protocols were approved by the Institutional Animal Care and Use Committees of Chonbuk National University (CBNU 2016-0030).
Biochemical analysis of serum
Serum rennin, angiotensin II, and aldosterone concentrations were analyzed using an ELISA kit (Rat Renin ELISA Kit, MyBioSource, San Diego, CA, USA; Angiotensin II ELISA Kit, and Aldosterone ELISA Kit, Enzo Life Sciences, Inc., Farmingdale, NY, USA).
RNA isolation and gene expression analyses
RNA was extracted from tissues using the Trizol method. The RNA synthesized to cDNA using high-capacity cDNA Reverse Transcription Kit (Applied Biosystem, Vilnius, Lithuania) with reverse transcription PCR (MWG Biotech Inc., Ebersberg, Germany). The cDNA analyzed as relative gene (Table 1) expression using SYBR Green (TOYOBO, Osaka, Japan) and Real-Time PCR (Applied Biosystems, Foster City, CA, USA).
Urine and feces analysis
Na+, K+, and Ca2+ concentrations in urine and feces were measured using inductively coupled plasma-mass spectroscopy (ICP-MS; 7500A, Agilent Technologies, Germantown, MD, USA). Samples were analyzed in the Center for University-Wide Research Facilities (CURF) at Chonbuk National University.
RESULTS
Systolic blood pressure
SBP in the NC group was higher than in the CT and GJ groups after oral administration of the diets (Fig. 2). There was a trend of decrease in SBP of GJ compared to other groups. SBP in the NC group increased 2 mmHg from 151.9 ± 13.65 mmHg to 153.47 ± 10.91 mmHg, while those in the CT and GJ groups decreased 10 mmHg from 150.87 ± 19.10 mmHg to 140.57 ± 11.10 mmHg and 19 mmHg from 155.3 ± 15.90 mmHg to 136.87 ± 11.57 mmHg, respectively Thus, on average, the SBP in the NC group was 13 mmHg higher than in the CT group and 14 mmHg in the GJ group during oral administration.
Excretion of Na+, K+, and Ca2+ and serum Na+ and K+ levels
Urine volume results from the 24 hour metabolic cage test were increased in GJ group compared to CT and NC, but body weight, food intake and water intake were not significantly different among groups (Table 2). However, ion contents in urine and feces were significantly different. In urine, Na+ was higher in the NC (6,828.67 ± 1,671.46 ppm) and GJ (6,206.0 ± 777.02 ppm) groups than in the CT group (1,060.83 ± 370.65 ppm); however, the results were not significantly different between the NC and GJ groups. The levels of K+ and Ca2+ ions were not significantly different among the 3 groups. Conversely, in feces, Na+ was not significantly different, but the levels of K+ and Ca2+ ions were decreased the GJ group than other groups (Table 3). Serum Na+ was higher in the NC group (137.00 ± 0.00 mEq/L) than in the GJ group (136.33 ± 1.37 mEq/L) and CT group (135.50 ± 0.84 mEq/L). In addition, K+ was highest in the GJ group (4.83 ± 0.74 mEq/L), followed by the CT group (4.40 ± 0.37 mEq/L) and the NC group (4.08 ± 0.12 mEq/L) (Table 4).
Renin, angiotensin II, and aldosterone levels in serum
Serum renin level was highest in the CT group (29.58 ± 9.91 pg/mL), followed by the NC group (25.2 ± 5.58 pg/mL) and GJ group (17.23 ± 2.50 pg/mL). Aldosterone level was highest in the NC group (877.73 ± 274.76 pg/mL), followed by the CT group (696.38 ± 134.52 pg/mL) and GJ group (546.15 ± 251.59 pg/mL). The serum rennin and aldosterone levels were lowest in the GJ group. However, serum angiotensin II level was not significantly different among the groups (Fig. 3).
Gene expression in kidney
Renin mRNA expression was relatively high in the CT group (1.03 ± 0.32), followed by the NC group (0.60 ± 0.30) and the GJ group (0.34 ± 0.08). Mineralocorticoid receptor (MR) expression was significantly lower in the GJ group compared with the other groups (CT, 1.00 ± 0.13; NC, 0.70 ± 0.16; GJ, 0.43 ± 0.07). Similarly, angiotensin II type 1 receptor (AT1 receptor) gene expression was higher in the CT group (1.02 ± 0.25) than in the NC group (0.65 ± 0.04) and GJ group (0.48 ± 0.07); expression in the NC and GJ groups was not significantly different. However, angiotensin I converting enzyme (ACE) gene expression was not significantly different among the groups (Fig. 4).
Na+ transporter gene expression showed similar results in urine and feces based on ion analysis. The GJ group was lower in Na+/K+ ATPaseα1 (NKAα1), Na+/H+ exchanger 3 (NHE3), Na+/HCO3- (bicarbonate) co-exchanger (NBC) and carbonic anhydrases II (CAII) than the NC group. The CT group also showed lower expression than the NC group in most of the sodium transporters except NKAα1. However, Na-Ca exchanger (NCX) gene expression was not significantly different among the groups (Fig. 5).
DISCUSSION
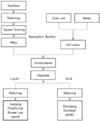
Dietary sodium can increase blood Na+ concentration and extracellular fluid volume. Therefore, sodium intake is closely associated with blood pressure [5614]. Additionally, a long-term high-salt diet induced renal injury and alteration of the renal cytokine genes followed by hypertension in normal rats [15]. In present study, blood pressure was significantly different in ganjang and sodium water despite the same concentration of NaCl in the groups. Therefore, ganjang may affect the RAAS differently compared to salt water and have a weaker negative effect on kidneys. High consumption of salt-fermented vegetables (kimchi) has not shown to be associated with increased risk of hypertension in adults [16]. Fermented rice bran decreased SBP in stroke-prone spontaneously hypertensive rats [17]. The metabolic syndrome related hypertension was improved by fermented red ginseng [18]. Microorganisms, dietary fiber, and phenolic compounds can be effective in the prevention of lifestyle-related diseases. The fermented milk and its peptide by lactic acid bacteria had hypotensive effect by inhibiting ACE activity, hydrolysis peptides, and vasodilator [1920].
In present study, we investigated whether the same concentrations of salts in ganjang and salt itself may have different effects in hypertension-induced rats. As shown in Table 2, the amounts of sodium and potassium intake were different in NC and GJ groups supplemented with same concentration salts. Interestingly, NC had 92 mg/kg BW/day more sodium intake than GJ group. Reduced intake of sodium and increased intake of potassium can be important factors in preventing hypertension [21]. Therefore, the reduced sodium contents in ganjang compared to table salt may be responsible for the decreased blood pressure in GJ group.
Based on the results of ion contents in urine and feces, Na+ ion excretion was lower in CT group urine, and K+ and Ca2+ ion excretion were significantly higher in CT group feces. K+ and Ca2+ ions are exchanged ions during Na+ reabsorption, indicating high Na+ reabsorption activity in the CT group. Na+ ion excretion in feces was not significantly different between the GJ and NC groups, although K+ and Ca2+ ion excretion were significantly higher in NC group feces, indicating higher Na+ reabsorption activity in the NC group compared with the GJ group. K+ and Ca2+ were exchanged Na+ by NKAα1 and NCX that was controlled by RAAS. NKAα1 has the central role in sodium reabsorption [22] and Ca2+ entry mode of NCX is triggered by the salt-dependent hypertension [23]. Additionally, it was reported to urinary sodium excretion is significantly associated with blood pressure [24]. The sodium excretion in the hypertensive people was higher than normal people [25]. Although serum Na+ was increased in GJ group than CT group, but urinary volume significantly increased the GJ group compared to CT group. The lower RAAS activity and increased excretion of Na+ in GJ group can improve the blood pressure.
With regard to serum hormone concentrations, renin level was lower in the GJ group than in the NC group, although SBP was lower in the CT group. This result is considered because renin is activated when plasma sodium level is low and blood flow is decreased [26]. Renin was activated and catalyzed angiotensinogen into angiotensin I when the prorenin form bound to the kidney renin receptor. Additionally, the RAAS was instigated by rennin binding with rennin receptor [27]. A high concentration of renin was not correlated with high enzymatic activity. However, similar tendencies of renal rennin mRNA expression, rennin secretion, and plasma rennin activity [28]. The renin mRNA abundance was tended to be suppressed by the high-NaCl diet [29]. Renin and renal renin mRNA levels were lower in the NC and GJ groups than the CT group, although not significantly. We hypothesized that a significant difference occurred when catalyzing from a high angiotensinogen level to angiotensin I and high angiotensin II level. However, ACE mRNA and angiotensin II levels were not significantly different. The ACE mRNA and ACE activity showed similar tendencies [3]. However, the aldosterone level was not significantly higher in the NC group than in the CT group. In contrast, angiotensin receptor type 1 (receptor of angiotensin II) was significantly higher in the CT group than in the NC and GJ groups. Angiotensin II activates AT receptors; AT1 receptors mainly mediate the stimulation of aldosterone secretion and angiotensin II, but type 2 receptors (AT2 receptors) also activate stimulation of aldosterone secretion by local release of catecholamines [30]. The activated AT2 receptor was considered a potential variable for aldosterone secretion. The aldosterone increased the expression of MR and AT1 receptors, thus affecting not only angiotensin II, but also aldosterone levels [31].
Aldosterone regulates the transfer of Na+ and K+ in distal nephrons and stimulates expression of transporters by binding to MR, a ligand-dependent transcription factor [32]. The mRNA expression of MR was higher in the CT group than in the NC group, and that of the NC group was higher than that of the GJ group. These results indicate that MR is affected by aldosterone and AT1 receptor expression [33]. The gene expression of MR and AT1 receptors was similar to their protein expression [3435]. Angiotensin II and aldosterone stimulate Na+ reabsorption transporter expression through AT1 receptor and MR [323637].
Na+ reabsorption transporters were directly/indirectly regulated by AT1 receptor and MR. NKAα1 was significantly higher in the CT group than in the NC group and was significantly lower in the GJ group than in the NC group. Similar results were observed with the AT1 receptor and MR gene expression. NBC and NHE3 were also regulated by the AT1 receptor and interacted with CAII. These results show similar tendencies with CAII gene expression. NCX was not significantly different among the groups, but the results showed a similar tendency to the Ca2+ excretion in feces. To conclude, under prolonged high sodium intake, renin, aldosterone, AT1 receptor, and MR were similarly and/or more activated compared to conditions of normal sodium intake. The Na+ transporters increased, enhancing Na+ reabsorption and causing hypertension. However, in the case of ganjang, when intake with same sodium concentration compare to high sodium intake group (NC group), renin, aldosterone, and MR were significantly lower than in the NC group. The Na+ transporters in the GJ decreased, which decreased K+ and Ca2+ excretion in feces, resulting in decreased Na+ reabsorption compared with the NC group. Further study is warranted to investigate the bioactive component which is responsible for antihypertension effects of ganjang and its underlying mechanisms. Our results suggest that consumption of ganjang may have lower risk of hypertension than table salt (Fig. 6).









 XML Download
XML Download