

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Chemokines (CCs) play a variety of roles in intercellular signaling, particularly in the recruitment of leukocytes to inflammation sites [1]. Monocyte chemotactic protein-1 (MCP-1), also referred to as chemokine ligand 2 (CCL2), promotes transmigration and emigration of circulating monocytes into tissues, and exerts various effect on monocytes, including the induction of superoxide anions and expression of various proinflammatory genes [2345]. MCP-1 levels are elevated in the plasma of older persons in the apparent absence of cardiovascular disease [6]. Single nucleotide polymorphism (SNP) in the MCP-1 promoter region has been shown to affect gene transcription [7]. The -2518 G/A polymorphism in the MCP-1 is associated with inflammatory related diseases, including rheumatoid arthritis, and diabetic retinopathy [89]. It has also been reported that the G/G genotype of -2518 MCP-1 was associated with susceptibility to cardiovascular disease (CVD) and was identified as a genetic risk factor for severe coronary artery disease [10].
Dietary factors may influence gene expression [111213]. One case control study [11] reported a nutrigenetic interaction between dietary vitamin E and thioredoxin and catechol-Omethyltransferase gene polymorphisms on waist circumference. Dietary factors may also alter the risk of inflammatory disease. Many studies have reported that dietary factors such as fish oil, broccoli, and probiotics were associated with anti-inflammatory effects [141516]. However, clinical trial outcomes are mixed, which may be due to an interaction between genetic background and dietary intake. Gene variation may also influence dietary requirements, e.g. the new field of nutritional genomics, and raises the possibility of individual nutritional management for optimal health and disease prevention on the basis of an individual genome. Multiple studies have already shown significant gene × diet interactions. Ortega-Azorín et al. [17] reported that the association of fat mass and obesity associated (FTO) gene polymorphism with type 2 diabetes depends on diet, and that adherence to the Mediterranean diet counteracts the genetic predisposition. Serrano et al. [18] showed that metabolic response to 25-hydroxyvitamin D and soybean supplementation was dependent on vitamin D receptor GG genotype. Giacconi et al. [19] showed the effect of zinc transporter protein 2 Gln/Arg/Leu polymorphism on proinflammatory mediators and zinc homeostasis in the elderly population. Park et al. [20] showed gene-nutrient interactions between transient receptor potential vanilloid-1 variants re161364 and preference for oily taste, intake of oily foods, and fat intake. Thus supplements may have different effects depending on genetic predisposition. However, little is known about the association of MCP-1 polymorphism and dietary factors as nutrients or supplements in the Korean population.
Spirulina is a cyanobacterial blue-green algae, commonly used as a dietary supplement [21]. Several studies have demonstrated spirulina's beneficial effects and therapeutic functions [2223]. Other studies have shown that spirulina has beneficial effects on the treatment of malnutrition and other pathologies such as obesity, hypercholesterolemia, hypertension, and diabetes mellitus [242526]. Studies have reported spirulina was associated with modulation of the host immune system. Hayashi et al. [27] first published detailed studies on the immunomodulatory properties of dietary spirulina in mice. Dietary spirulina enhanced responses of phagocytic activity responses, interleukin (IL) -1b expression, and tumor necrosis factor (TNF)-α genes in carp [28]. Spirulina treated chicken macrophages showed increased spreading and vacuolization with minimal cytotoxicity [29]. In humans, spirulina increases natural killer cell cytotoxicity [30] and ameliorates immunosenescence in older subjects [31]. Spirulina supplementation produced a significant rise in plasma IL-2 level, and a significant reduction in IL-6 level in health elderly [32]. These results suggest that spirulina somehow stimulates the immune system.
From the perspective of personalized nutrition, the effect of spirulina on immune function is different depending on the individual's obesity [33]. No studies have investigated the effect of MCP-1 polymorphism on the effect of spirulina supplementation on immune function. Therefore, the aim of the current study was to explore interactions between spirulina supplementation and MCP-1 -2518 G/A polymorphism on immunity in Korean elderly.
SUBJECTS AND METHODS
Subjects and experimental design
Subjects were recruited through an advertisement in local newspapers (Dec 2005 to June 2006). The subjects were first interviewed by telephone for screening. Exclusion criteria were: current user of vitamin supplements, current drug-user for inflammatory disease (e.g. Crohn disease, rheumatoid arthritis), dyslipidemia, hypertension, concurrent or recent participant in any other intervention study.
The study protocol was approved by the Institutional Review Board of Ewha Women's University (ECT 109-02-01), and written informed consent was obtained from all study subjects. A randomized, controlled, double blinded, parallel group design study was performed for 78 healthy older adult subject (43 male, 35 female). Subjects consumed spirulina or placebo, 8 g per day, for 16 consecutive weeks. At the first visit of the study, blood was drawn after a minimum of 12 h fasting, defined as the baseline. Blood samples were taken again at the end of the study period. During the intervention period, subjects were asked to keep their usual diet, and prohibited to take any functional food or supplements. Both spirulina and placebo (100% starch) were provided by Earth Spirulina Biotech (Seoul, Korea). Spirulina and placebo were suppled every 2 weeks and compliance was confirmed by telephone twice a week.
Food consumption and anthropometric measures
The elderly subjects were interviewed individually to obtain data on food consumption and general characteristics. Food consumption was assessed using the 24 hour recall method. Food intake data was analyzed using CAN-pro 4.0 software [34].
Standing height was measured using an anthropometer (Seca 213, Seca Inc. Birmingham, UK). Body weight and composition [body fat (kg), body fat (%), and lean body mass (kg)] were measured using INBODY 2.0 (Biospace co, Seoul, Korea) with subjects wearing light clothing without shoes or socks.
Blood collection and analysis
Blood samples collected before and after supplementation from overnight were taken on the beginning day (pretest) and 16th week (posttest) of the study period. Fasting blood samples were obtained and sera were kept at 80℃ until measurement. Total cholesterol and triglyceride level were measured using an autoanalyzer (Ekachem DTSC module, Johnson & Johnson, New Brunswick, NJ, USA). HDL cholesterol level was determined by autoanalyzer after treating UC infranatant with phosphotungstic acid Mg. LDL cholesterol was calculated as described by Friedewald [35].
Plasma MCP-1, IL-2, IL-6, and TNF-α concentrations were determined by enzyme linked immunosorbent assay (ELISA) (Quantikine, R&D Systems Inc., Minneapolis, MN, USA) using an ELISA reader (Spectra Max 340, Molecular Devices, CA, USA). Radial immunodiffusion plates (Norpartigen, Behring Co., Marburg, Germany) were used to measure Ig and C3 level. Assays were performed as recommended by the manufactures.
The proliferation capacity of lymphocytes was analyzed by the Mosmann method [36]. Fresh blood samples treated with heparin were diluted in phosphate buffered saline (PBS) 1:1, and then cells were separated in 3 mL Ficoll-paque gradient (lymphocyte isolation solution, Histopaque 1077, Sigma, MO, USA) by centrifuge at 1,000 g for 30 min. The isolated peripheral blood mononuclear cells (PBMCs) were seeded at 2 × 105 cell per well in a 96-well plate. PBMCs were treated with concanavalin A (ConA, Sigma, MO, USA, 0.5 µg/10 µl), phytohemagglutinin (PHA, ThermoFisher Scientific co., MA, USA, 0.5 µg/10 µl), and lipopolysaccharide (LPS, Sigma, MO, USA, 0.5 µg/10 µl), and then incubated for 68 h in a CO2 incubator. Viable cell count was estimated by 3-[4, 5-dimethylthizol-2-yl]-2,5-diphenyl tetrazoliumbromide (MTT, Sigma, MO, USA) assay. Peripheral blood lymphocytes proliferation rate (LPR) was estimated by the stimulation index, SI = absorbance of wells with mitogens / absorbance of wells without mitogen. All tests were run in triplicate.
DNA extraction and analysis of the MCP-1 -2518 G/A polymorphism
All subjects were genotyped for MCP-1 polymorphism (-2518 G/A) (dbSNP: rs1024611). DNA was extracted from whole blood (QIAamp DNA Blood Kit, Qiagen, Hilden, Germany), and a 649 bp segment of MCP-1 gene including polymorphism sites was amplified by polymerase chain reaction (forward primer: 5′-AGT CCA ACC AAG GTT TGT GC-3′, and reverse primer 5′-TCA TGC TTC GGG TTT TCT CT-3′). The presence of MCP-1 polymorphism was determined by restriction fragment length polymorphism (RFLP) assay using PvuII enzyme.
Statistical analysis
Statistical analysis was conducted using SAS Statistics version 9.4 (SAS Institute, NC, USA). Data are presented as mean ± standard error (SE) for placebo and spirulina groups by genotype. Paired t-test was used to analyze mean differences for all measured parameters between baseline and week 16. Repeated measure ANOVA was conducted to examine treatment effects.
RESULTS
Subjects' baseline characteristics
Seventy-eight healthy subjects were divided into placebo (p, n = 37) and spirulina groups (s, n = 41). The proportions of A/A, G/A, and G/G genotypes were 17% (n = 14, p/s = 5/9), 56% (n = 44, p/s = 19/25), and 25% (n = 20, p/s = 13/7), respectively. Selected baseline characteristics are summarized in Table 1. Average subject age was 66.0 years. There were no significant differences in age, BMI, lipid profile, and dietary intake between 2 groups within the same genotype. Three day dietary records showed no significant intergroup and intragroup differences in mean daily intakes of energy, protein, carbohydrates, total fat, vitamin A, vitamin C, and vitamin E.
Spirulina effects on plasma cytokine based on genotype
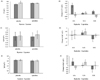
Plasma MCP-1 level did not differ significantly among the intervention group at baseline (placebo 80.9 pg/dL, spirulina 77.8 pg/dL) and after supplementation (placebo 77.2 pg/dL, spirulina 74.3 pg/dL) (Fig. 1A). When the plasma MCP-1 level of subjects was analyzed by genotype, MCP-1 level was significantly increased in the placebo group (P < 0.05) with AA genotype, whereas there was no change in the spirulina group (Fig. 1B). No significant difference in the IL-6 level was observed for the intervention group or genotype groups (Figs. 1C, 1D). Plasma TNF-α level was significantly decreased after spirulina supplementation for all subjects (Fig. 1E, P < 0.05), but this significance was not observed for genotype subgroups (Fig. 1F). IL-2 level was significantly increased in the spirulina group after supplementation (Fig. 1G), and the significant was observed in all genotypes (Fig. 1H).
Table 2 shows the cytokine changes after spirulina intervention between the A/A genotype (n = 14) and G/A + G/G MCP-1 genotypes (n = 64). There were significant differences in plasma MCP-1 levels for MCP-1 genotype and MCP-1 genotype × spirulina treatment (P < 0.01, P < 0.05), whereas there were no significant differences in IL-6 levels. The level of IL-2 increased in the spirulina group (P < 0.05) but not in the placebo group, irrespectively of MCP-1 genotype. Although TNF-α level was significantly decreased (P < 0.05) in both groups with G carriers, there was no significant difference by genotype after supplementation.
Spirulina effects on complement and Ig based on genotype
C3 level significantly increased after placebo supplementation for 16 weeks (Fig. 2A, P < 0.05). However, C3 level was not changed in all groups after spirulina supplementation, but only for subjects with A genotype (P < 0.05) (Fig. 2B). There were no significant differences of IgA and IgG levels in all genotypes after spirulina supplementation (Figs. 2D and 2F).
Table 3 shows C3 and Ig changes after placebo or spirulina supplementation for the A and G genotype subgroups. There was significant spirulina supplementation effect on C3 level (P < 0.01), and also MCP-1 genotype × treatment for C3 (P < 0.05).
Spirulina effects on lymphocyte proliferation rate based on genotype
Lymphocyte proliferation rate showed no significant difference for all subjects after spirulina or placebo supplementation (Figs. 3A, 3C, 3E). Changes in stimulation index of mitogen induced LPR from PBMC are shown in Figs. 3B, 3D, and 3F by genotype. PHA induced SI was significantly changed for A/A and G/A genotypes (Fig. 3D, P < 0.05). LPS induced SI was significantly changed only for the A/A genotype (Fig. 3F, P < 0.05).
A significant difference of intervention between the A and G genotype subgroups appeared in PHA and LPS induced proliferation rate with A carrier (Table 4). Mitogen induced proliferation of lymphocyte was not significantly changed for both G carrier groups. A significant supplementary effect was observed for the case of PHA induced SI, regardless of genotype (P < 0.01).
DISCUSSION
MCP-1 is a small cytokine that belongs to the CC chemokine family [37]. The concentration gradient of MCP-1 is responsible for the movement of mononuclear cells to inflammation sites [38]. MCP-1 is produced by a variety of cell types, including immune and non-immune cells [3473940], in response to stimulation by a broad range of mediators, including cytokines, growth factors and mitogens [4]. MCP-1 recruits monocytes, memory T cells, and dendritic cells to inflammation sites produced by either tissue injury or infection [3841]. MCP-1 is also implicated in pathogeneses of several diseases characterized by monocytic infiltrates, such as psoriasis, rheumatoid arthritis, and atherosclerosis [40]. Epidemiological evidence suggests that MCP-1 plasma levels are associated with traditional risk factors for CVD, such as hyperlipidemia, hypertension, and diabetes [4243]. MCP-1 may also contribute to thrombotic aspects of advanced atherosclerotic lesions, a late complication of atherosclerosis responsible for myocardial infarctions and strokes [37]. MCP-1 expression is up-regulated in human atherosclerotic plaques [37], suggesting that genetic factors leading to MCP-1 production deregulation could affect development of atherosclerosis and associated diseases.
The human MCP-1 gene is located on chromosome 17q11.2-q21.1 [37]. Several MCP-1 SNPs, such as -1811 A/G, -2136 A/T, -2518 A/G, -2835 C/A, -927 G/, 764 C/G, and 3726 T/C have been identified [44]. Among these, MCP-1 -2518 A/G SNP, located in the MCP-1 regulatory region, has been most frequently studied for its relevance to diet and disease. Szalai et al. [44] found that MCP-1 -2518*G allele homozygotes frequency was significantly higher in coronary artery disease (CAD) patients than in healthy controls. The higher MCP-1 -2518*G homozygotes prevalence was observed in patients with history of stroke [45] and myocardial infarction [46]. Angeles-Martinez showed that MCP-1 polymorphisms were associated with premature coronary artery disease risk [47]. Other studies [4849] have reported MCP-1 polymorphism association with disease related inflammation susceptibility.
Several studies have investigated the relationship between MCP-1 -2518A/G polymorphism and CVD risk in various populations worldwide [50515253]. However, the results are inconsistent, and MCP-1 -2518 A/G polymorphism appears to be ethnically different. Caucasian or African subjects are less likely to express MCP-1 -2518 G/G polymorphism than Asian or Mexican [7]. On the other hand, Caucasian or African subjects with MCP-1 -2518 G/G polymorphism show higher MCP-1 plasma levels and higher CVD risk, such as hypertension, than those with other genotypes [54]. In Chinese subjects, MCP-1 -2518 A/G polymorphism was not associated with CAD risk [50]. In a previous study, we reported that subjects with A/A genotype exhibited higher MCP-1 levels than other genotypes in Korean elderly, and had higher CVD risk [55]. These results suggest that MCP-1 concentration and CVD risk vary with MCP-1 polymorphism.
In the current study, the effects of spirulina supplementation were different depending on MCP-1 genotypes. Anti-inflammatory spirulina supplementation effects in all subjects were not significant, but the effect of intervention was significantly different when analyzed by genotype. For A/A genotype subjects, spirulina supplementation showed significant changes for plasma IL-2 level and PHA or LPS induced LPR. Placebo subjects showed significant MCP-1 increase, but those who took spirulina did not. Circulating MCP-1 levels are associated with CVD risk factors, since MCP-1 is expressed at high levels in atherosclerotic plaque [52535457]. IL-2 is a representative anti-inflammatory cytokine that decreases with aging [58].
C3 is a protein with a central role in the innate immune system, and C3 levels have an aging dependent increase in general. Several studies have reported that C3 was positively related to CVD [5960]. In the current study, placebo subjects showed significant C3 level increase after 16 weeks, but there was no significant difference in spirulina subjects. Subjects with A/A genotypes also had significantly more C3 compared with G/A and G/G genotypes when analyzed by genotype and placebo supplementation: A/A increased 32.8%, and G/A and G/G increased 7.5%. These results indicate that the degree of C3 increase depends on the MCP-1 genotype and that C3 increase was suppressed when spirulina was ingested.
Mitogens, such as PHA, ConA, and LPS induced stimulation index, are generally reduced by aging [61]. The current study shows that lymphocyte proliferation was improved after spirulina supplementation. Subjects who consumed spirulina for 16 weeks showed significantly increased PHA and LPS induced stimulation index. Significantly, the PHA and LPS induced stimulation index also increased in A/A genotype subjects.
Thus, spirulina supplementation shows anti-inflammatory effects, such as decreasing MCP-1 level and increasing IL-2 level in healthy subjects, and improved LPS and PHA induced mitogenesis. Theses anti-inflammatory effects were larger for A/A genotype subjects. Therefore, spirulina may be considered beneficial for anti-inflammatory capacity, particularly for A/A genotype subjects.
In conclusion, functional food supplementation has different effects by genotype. Spirulina supplementation affects anti-inflammatory capacity and may help immune function. Although the population size of the current study was small, the study confirmed that supplement response and anti-inflammatory profiles were different by MCP-1 genotype. These results suggest that personalized management could be advantageous for different genotypes. While the current study is insufficient to make specific personalized nutritional recommendations based on genetic information, the results provide a valuable basis for further study to investigate the relationship between MCP-1 polymorphism and diet. Careful attention should be paid for subsequent studies to include sufficient subjects for each phenotype and genotype with appropriate statistical power.






 XML Download
XML Download