

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Obesity is associated with insulin resistance [12], which is defined as a reduction in the ability of insulin to activate its signaling pathway in the main target organs [34]. Obesity-caused dysfunction of adipose tissue is strongly associated with increased incidence of type 2 diabetes (T2D) and cardiovascular diseases [5], because adipose tissue plays a critical role in regulation of insulin action, and glucose and lipid metabolism [67]. Insulin increases glucose uptake in adipose tissue and muscle, and inhibits glucose output from the liver. Binding of insulin to the receptors on insulin responsive cells, leads to phosphorylation of the tyrosine residues on the insulin receptor substrates (IRSs) by tyrosine kinase. Phosphorylated IRSs (p-IRSs) serve as docking proteins for SH2-containing enzymes, leading to the activation of signaling cascades that result in Akt phosphorylation. The phosphorylation of Akt activates glucose transporter type 4 (GLUT4) and induces glycogen synthesis [89]. The activation of glucose transport via GLUT is the rate-limiting step during glucose uptake [10]. GLUT4, predominantly expressed in adipose tissues and muscle, plays a crucial role in glucose homeostasis [11]. The inhibition of GLUT4 expression in adipose tissue results in insulin resistance [12].
The most commonly used drugs show adverse effects and usually high doses are required to improve insulin resistance [13]. Thiazolidinediones (TZDs), a class of peroxisome proliferator-activated receptor (PPAR)-γ agonists for treating T2D, promote adipogenesis and increase insulin sensitivity [1415]. However, the clinical use of TZDs has several adverse effects such as weight gain, fluid retention, congestive heart failure, and bone fracture [1617]. Furthermore, TZDs-induced weight gain will deteriorate insulin resistance. Therefore, there is an urgent need for novel effective drugs to regulate adipocyte function, especially to protect obese patients against insulin resistance and other obesity-related diseases. For this reason, research efforts are directed toward development of safe and cost-effective natural alternatives to conventional drugs commonly used to improve insulin sensitivity [1819].
Pear is consumed as a fresh fruit and fruit products worldwide. However, pomace, the solid residue produced during manufacturing, is discarded, which causes environmental pollutions and economic waste. Pear pomace, which is composed of peels, pulps, stems, cores, and seeds, is known to contain lignocellulose-rich materials [20]. Pear has important dietary components including polyphenols and triterpenes, which exhibit antioxidant and anti-inflammatory actions [21]. Furthermore, polyphenol-rich plant extracts have inhibitory actions on α-glucosidase and α-amylase enzyme activities in T2D [222324]. The anti-diabetic activity of pear peel and pulp extracts through inhibition of α-glucosidase activity has been reported [25].
However, little has been reported about the insulin signaling pathway as an underlying mechanism of pear induced insulin sensitivity improvement. Therefore, further studies are required to establish the positive effects and the underlying mechanisms of action of pear pomace on insulin sensitivity. In this study, we conducted both cell-based assay (in vitro) and animal study (in vivo) with 50% ethanol extract of pear pomace (PPE) to explore the role of pear pomace in improving insulin sensitivity.
MATERIALS AND METHODS
Preparation of extracts from pears
Pears (Pyrus pyrifolia Niitaka) were washed, de-seeded, and then juices were extracted. Pear pomace, the juice extract cake obtained from pears, was freeze-dried and grinded to powder. Powdered pear pomace (100 g) was then extracted with 2 L of 50% ethanol for 24 h at room temperature. The extract was concentrated in a rotary vacuum evaporator, freeze-dried to a power, and then stored in a deep freezer (-70℃). The yield of pear pomace ethanol extract (PPE) obtained was 27.5%.
Cell culture, cell viability and differentiation of 3T3-L1 preadipocytes
Cell culture and assessments for viability and differentiation were performed as described previously [5].
Lipid accumulation, MCP-1 secretion, and glucose uptake in 3T3-L1 cells
Oil Red O (ORO) staining was performed on day 8. To measure the lipid accumulation in 3T3-L1 preadipocytes, cells were fixed with 10% formalin and stained with ORO. ORO dye was extracted by isopropanol to quantify relative TG accumulation [5]. The level of monocyte chemotactic protein-1 (MCP-1) in the supernatants were detected by using ELISA kit (Thermo Fisher Scientific Co., Rockford, IL, USA) according to the manufacture's protocol. The standard curve of MCP-1 was 16-1000 pg/mL. All standards and samples were assayed in triplicate. Cellular glucose uptake was quantified by the 2-[N-(7-notrobenz-2-oxa-1, 3-diazol-4-yl) Amino]-2-Deoxyglucose (2-NBDG, Invitrogen, Carlsbad, CA, USA) assay. 3T3-L1 cells were seeded in 96 well black plates and differentiated into mature adipocytes as reported previously [5]. On day 6, the cells treated with PPE. After 24 h, treatment medium was removed and added with 200 µL 2-NBDG solution for 30 min. Then, cells were washed twice with PBS, the cells were measured with a VICTOR™ Multilabel Counter (PerkinElmer, USA), set at an excitation wavelength of 465 nm and an emission wavelength of 540 nm [26].
Western blot analysis from 3T3-L1 cells
Mature adipocytes were treated with PPE for 24 h. Medium was removed and replaced with DMEM containing 10 nM insulin for 30 min. After washing with PBS, cells were collected in lysis buffer, followed by centrifugation at 13,000 rpm for 20 min to collect the supernatant. Protein concentrations were determined by the Bradford (Sigma, St. Louis, MO, USA) method and equal amounts of protein (25 µg for each) were separated in 10% SDS-PAGE gels and transferred onto a nitrocellulose membrane (Millipore, Billerica, MA, USA). The membrane was blocked for 1 h at 4℃ with 5% nonfat milk in TBS-T buffer [10 mM Tris-HCl (pH 6.8), 100 mM NaCl, and 0.1% Tween 20]. The membranes were then incubated with primary antibodies [p-IRS-1 (Ser307) 1:1000, IRS-1 1:1000, Akt 1:1000, p-Akt (Ser473) 1:1000, and GLUT4 1:1000; Cell Signaling Technology, Beverly, MA, USA; p-IRS-1 (Tyr632) 1:1000 and β-actin 1:1000; Santa Cruz Biotechnology Inc., Dallas, Tex, USA] overnight at 4℃ and then with horseradish peroxidase conjugated secondary antibody [goat anti-rabbit or goat anti-mouse (1:2000), Jackson ImmunoResearch Inc, West Grove, PA, USA] for 1 h. Bands were detected by chemiluminescent substrate (IMGENEX ,San Diego, CA, USA), and the protein expression was quantified using UVP (imaging system for chemiluminescent Western blot, Upland, CA, USA) and Vision Works TMLS (Analysis Software, Upland, CA, USA).
Animals
The study protocol was approved by the Animal Care and Use Committee of Mokpo National University (MNU-IACUC-2014-006). Five week old male C57BL/6 mice were obtained from Central Lab. Animal, Inc. (Seoul, Republic of Korea). All mice were maintained in a climate-controlled room (temperature and relative humidity maintained at 22 ± 2℃, 50 ± 10%, respectively) under 12 h light/dark cycle, and provided with diet and water ad libitum. At 6 weeks of age, the mice were randomly divided into three groups (n = 10/group) and fed a high-fat diet (HFD; 60% kcal fat) for 8 weeks. The mice were administrated with PPE (200 or 400 mg/kg body weight dissolved in distilled water) by oral gavage once per a day. Change in body weight of all mice was monitored weekly. At week 8, blood was collected after 8 h of fasting and serum was separated. Tissues, including liver and abdominal adipose tissue were removed, washed with phosphate-buffered saline (PBS), and weighed. Adipose tissue was snap-frozen immediately in liquid nitrogen and stored at -70℃ for further analysis.
Measurement of blood biochemical parameters
Fasting blood glucose concentration was determined using a commercial kit (Allmedicus, gyeonggi-do, Republic of Korea) at week 8. Fasting serum insulin (Shibayagi Co. Ltd, Gumma Pref, Japan) was also measured using a commercial kit. Serum lipids including triglyceride (TG), total cholesterol (TC), and LDL-cholesterol were determined using an Automated Chemistry Analyzer (Beckman Coulter Inc, Brea, Cal, USA).
Western blot analysis in mice
Snap frozen adipose tissues were homogenized in ice-cold radioimmune precipitation assay (RIPA) buffer (Thermo Fisher Scientific Co., Rockford, IL, USA) with 1% protease inhibitors cocktail and phosphatase inhibitor. The cell lysates centrifuged at 13,000 rpm for 20 min to collect the supernatant. The method for immunoblotting was same as described in cell-based assay.
Statistical analysis
Statistical analysis was performed by one-way ANOVA followed by post hoc Duncan's multiple comparison test. Data are presented as the mean ± SE for three independently performed experiments. P-value < 0.05 was considered as statistically significant. Statistical analysis of all data was conducted using SPSS version 23.0.
RESULTS
Effects of PPE on the viability and the differentiation of 3T3-L1 preadipocytes
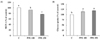
3T3-L1 cells were incubated with PPE for 24 h before MTT assay. MTT assay showed that the PPE did not affect cell viability at concentrations of 100 and 250 µg/mL (Fig. 1A). Changes in TG accumulation in response to PPE treatment were determined at 8 day. The PPE did not affect TG accumulation (Fig. 1B).
Effects of PPE on MCP-1 secretion and the glucose uptake in 3T3-L1 cells
MCP-1 secretion and glucose uptake in 3T3-L1 cells treated with PPE were determined. As shown in Fig. 2, PPE significantly decreased MCP-1 secretion (A) and increased glucose uptake (B) compared with control.
Effects of PPE on the insulin signaling pathway in 3T3-L1 cells
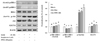
To determine the underlying mechanism of action of PPE in improving insulin sensitivity, we determined the expression of proteins involved in insulin signaling pathway such as IRS-1, p-IRS-1 (Tyr632/Ser307), Akt, p-Akt (Ser473), and GLUT4 (Fig. 3). PPE significantly increased the expression of proteins such as p-IRS-1 (Tyr632) and p-Akt, and the increasing effect of PPE was bigger than that of rosiglitazone (1 µM). PPE also increased the expression of GLUT4 to the same level as rosiglitazone did. Contrary to our expectation, PPE did not decrease the expression of p-IRS-1 (Ser307) in 3T3-L1 cells activated by insulin (Fig. 3).
Effects of PPE on the account of weight gain and organ weight in high-fat diet-fed mice.
The body weight and the amount of weight gain of the mice treated with 400 mg/kg PPE significantly decreased in comparison to those of the mice in the control group (Fig. 4A and B). PPE at a dose of 400 mg/kg/day decreased intra-abdominal adipose tissue weight (Fig. 4C). The relative liver weight was calculated by dividing the weight of the liver by the body weight (liver weight/body weight). The PPE did not reduce relative liver weight (Fig. 4D). Food efficacy ratio was calculated as the weight gain/food intake during the experimental period. PPE at a dose of 400 mg/kg/day tended to reduce food efficacy ratio, but PPE-induced reduction was not statistically significant (Fig. 4E). Weight gain and adipose tissue weight in mice treated with PPE at a dose of 200 mg/kg/day tend to be lower than those in control mice, but the differences were not statistically significant.
Effects of PPE on insulin resistance and levels of serum lipids in high-fat diet-fed mice
PPE at a dose of 400 mg/kg/day reduced fasting blood insulin, however, the difference was not statistically significant (Fig. 5B). Nevertheless, 400 mg/kg/day of PPE significantly decreased HOMA-IR (Fig. 5C). Serum triglyceride (TG) and LDL-cholesterol (LDL-C) concentrations in mice treated with PPE at a dose of 400 mg/kg/day significantly decreased (Fig. 5D and F). However, no significant difference was observed in serum total cholesterol (TC) when compared with that of control group mice (Fig. 5E).
Effects of PPE on the insulin signaling pathway of adipose tissue in high-fat diet-fed mice
To identify the underlying action mechanisms of PPE in improving insulin sensitivity in high-fat diet-fed animal model, we determined the expression of proteins involved in insulin signaling pathway such as IRS-1, p-IRS-1 (Tyr632/Ser307), and GLUT4 in the adipose tissue (Fig. 6). Similar to 3T3-L1 cells, mice treated with PPE showed a significant increase in the expression of proteins such as p-IRS-1 (Tyr632), and GLUT4. Furthermore, PPE treated mice showed a decrease in the protein expression of p-IRS-1 (Ser307).
DISCUSSION
Owing to the adverse effects of commonly used anti-diabetic drugs, the current research trend is to identify and develop newer and safer alternatives for management of diabetes and insulin resistance. In the present study, we demonstrate the insulin sensitivity improving effects of PPE via activation of insulin signaling pathway in 3T3-L1 cells and high-fat diet-fed animals.
Adipose tissue plays an important role in the regulation of glucose and lipid metabolism, energy balance, inflammation, and insulin action [67]. In obesity, adipose tissue is the major site for increased macrophage infiltration. MCP-1 recruits macrophages into adipose tissue. Macrophages promote proinflammatory state and result in insulin resistance in adipocytes by inhibiting GLUT4 expression [1229]. In the present study, PPE decreased MCP-1 secretion and increased glucose uptake in 3T3-L1 cells (Fig. 2). However, both macrophages and adipocytes produce proinflammatory cytokines and these cytokines synergize and regulate each other [12]. In our experiment, we observed PPE-induced decrease in MCP-1 secretion in 3T3-L1 cells without coculturing with macrophages or treatment of TNF-δ. Therefore we need more researches to demonstrate the triggering effect of PPE on GLUT4 expression via a downregulation of MCP-1 secretion. The anti-oxidative and anti-inflammatory effects of pear fruit eaten with peal have been also reported [2125]. We also revealed that PPE reduced HOMA-IR in high-fat diet-fed animals, indicating that PPE can also improve insulin sensitivity in an in vivo model.
Several studies have been conducted on the insulin signaling pathway and revealed disruption of this insulin signaling cascade can induce insulin resistance [89101112]. Therefore, we can assume that agents that improve obesity-induced disruption of insulin signaling cascade can be developed as novel therapeutic agents. Thus, we determined whether PPE can improve insulin sensitivity through action on the insulin signaling pathway. In the present study, PPE increased tyrosine phosphorylation of IRS-1 thereby, increasing the ratio of p-IRS-1 (Tyr632)/IRS-1, which in turn increased phosphorylation of Akt in 3T3-L1 cells (Fig. 3). Furthermore, PPE increased GLUT4 expression in 3T3-L1 cells (Fig. 3). GLUT4 is the primary transporter for glucose uptake in adipose tissue [30]. Acquired insulin resistance of glucose uptake, observed in diabetes, could be explained by a variety of defects in GLUT4, including alteration in GLUT4 expression and translocation [31]. Decreased expression of GLUT4 and impaired responsiveness of GLUT4 to insulin were observed in adipocytes of obese and diabetic patients [3233]. Therefore, our results suggest that PPE improves insulin sensitivity through the activation of insulin signaling pathway in 3T3-L1 cells. The activation of insulin signaling pathway by PPE was also demonstrated in a high-fat diet-fed animal model (Fig. 6). High-fat diet is a nutritional condition that accounts for the high incidence of metabolic syndrome in the world [34]. In animal model, a high-fat diet feeding leads to an elevation of fasting blood glucose and insulin accompanied by an increased HOMA-IR index [35]. Fraulob et al. [36] demonstrated that C57BL/6 mice fed high-fat chow (60% calories from fat) distinctly displayed an increased HOMA-IR index that is compatible with the presence of insulin resistance in these animals. In the present study, similar to its effects on 3T3-L1 cells, high-fat diet-fed animals treated with PPE showed increases in p-IRS-1 (Tyr632)/IRS-1 in adipose tissue. Moreover, PPE also inhibited serine phosphorylation of IRS-1 in adipose tissue of high-fat diet-fed animals, which was hardly revealed in 3T3-L1 cells treated with PPE. Serine phosphorylation of IRS-1 inhibits the tyripsin phosphorylation of IRS-1 and disrupts insulin signaling cascade [37]. We could not determine the protein expression of p-Akt in the adipose tissue of high-fat fed animals, because Akt phosphorylation is significantly lower in adipose tissue than live tissue without insulin stimulation [3839]. PPE-induced increase in GLUT4 expression was also demonstrated in adipose tissue of high-fat diet-fed animals (Fig. 6). Therefore, we confirmed insulin sensitivity improving effect of PPE and demonstrate that PPE improves high-fat diet-induced insulin resistance through activation of insulin signaling pathway.
Obese individuals are assumed to ultimately develop diabetes, with insulin resistance occurring years before definite diagnosis. Therefore, agents that improve insulin resistance can prevent obese individuals from developing clinically overt diabetes. Weight gain is the one of the frequently occurring side effects of commonly used anti-diabetic drugs, indicating that these classes of drugs cannot protect obese patients against insulin resistance and diabetes. With respect to the metabolic effects of PPE, it should be noted that PPE did not affect lipid accumulation in 3T3-L1 cells (Fig. 1). Moreover, it decreased body weight and adipose tissue weight in high-fat diet-fed animals (Fig. 4). PPE also demonstrated hypolipidemic effects in high-fat diet-fed animals (Fig. 5). Further study need to elucidate two separate underlying mechanisms of PPE responsible for insulin sensitivity and adipogenesis. Some researchers have reported the therapeutic alternatives in the improvement of insulin resistance through enhancement of insulin signaling pathway without body weight gain [40]. They suggest the activation of AMPK [40], alteration of gut microbiota composition [35] and down-regulation of mTOR [41] as underlying mechanisms of anti-obesity effect.
In conclusion, PPE demonstrated insulin sensitivity improving effects via activation of insulin signaling pathway in 3T3-L1 cells (in vitro) and in high-fat diet-fed animal model (in vivo). Notably, the unique properties of PPE suggested that there might be two separate underlying mechanisms responsible for increased insulin sensitivity and adipogenesis.




 XML Download
XML Download