

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Immunopotentiation activity is a function that prevents the decrease of immunity accompanied by aging or diseases, and it is expected that it can be utilized in the development of new functional foods by investigating functions from commonly used foodstuffs and further separating and interpreting functional materials.
Yam (Sanyak) is a perennial herb (vine), which is native to China but has grown wild and usually seen in the mountains countrywide. The edible part is root which is collected at late autumn and then dried in the shades after peeling off the outer skin. Edible yam grown wild naturally or cultivated in the country can be divided into long yam with its roots deep in the ground, short yam with relatively shorter root tubers, and Chinese yam with long columnar roots, depending on the shape of tuberous roots, among which short yam was introduced from Japanese yam and cultivated, which has been convenient in harvesting and high in productivity and thus distributed for cultivation for foodstuffs and widely used for processing (Lee, 1982; Medicine Botany Society for the Research, 1993). Yam contains saponin, tannin, polyphenol, vitamins, sugars, amino acids, uronic acid, chelidonic acid, sitosterol, batasin, and dioscin in addition to general components, and thus has been used as an ingredient for nourishing, tonic, spleen invigorating, restorative, antidiarrheal medicines in the oriental medicine, in which the periderm of its tuberous root is removed and then dried as it is or steamed and then dried and has been called as 'Sanyak' since old times. Because of these health-oriented images of yam, it is recognized as an ingredient for oriental herb medicine or a material for functional food and thus its production and consumption have been increased and several agricultural cooperatives and companies have processed and sold yam as dried powder.
Several studies on yam were reported including studies on starch characteristics of yam (Choi et al., 1992; Kim et al., 1991; Lee et al., 1992) and studies on processing characteristics (Jung et al., 1996; Kwon et al., 1998; Lee et al., 1998). For studies on mucilage, a characteristic of yam, Han et al. (1990) investigated the degree of viscosity and components of mucilage of raw (fresh) yam and steamed-dried yam and reported that the degree of viscosity of steamed-dried yam was considerably lower than that of raw yam and the protein content was also low, but the mannose content was higher in steamed-dried yam. Lee et al. (1994) reported that mucilage could be separated if yam was freeze dried and ultra-fine milled into 5-30 µm size particles and then air classified at over 15,000 rpm. For in vitro experiments using yam mucilage, Ha et al. (1998) studied the heavy metals (Co, Cr, Cu) elimination capacity and angiotensin converting enzyme (ACE) inhibitory effect of yam mucilage and reported that since the adsorption capacity for heavy metals is one of important functions of polysaccharides, for example, the heavy metal adsorption capacity by chitosan and zooglan has already been reported, and the heavy metal adsorption capacity by yam mucilage was excellent and also the inhibitory effect on blood pressure increase would be expected because of ACE inhibitory action. In addition, some in vivo experiments including the study on the effect of yam on blood glucose in diabetics (Kwon et al., 2001a) and the study on the anti-mutagenicity and anti-complementary activity of yam by Lee (1996). Several foreign studies were performed on the components of yam mucilage, and Misaki et al. (1972) reported that major components of mucilage were protein (71.8%) and mannan (13.8%), and the mean degree of polymerization of mannan was 110. Tomoda et al. (1981) reported that mucilage was made of partially acetylated mannan and proteins containing some phosphorus, and its ratio was 1:2.2. According to Ohtani and Murakami (1991), mannan was made of about 90% of mannose and 10% of galactose and glucose. Tanoue and Simozono (1991) also reported that polysaccharides of yam were mainly mannose and some of arabinose, glucose, and galactose, and very small amount of xylose and rhamnose.
Physiological activities of yam have been known to be caused by mucilage, a peculiar characteristic of yam, but the efficacy of yam has mostly come from oriental medicine or folk remedy and few studies were performed scientifically on the application to immune system or its mechanism, except some recent studies on heavy metal elimination capacity and angiotensin converting enzyme inhibitory effect, blood glucose decrease, anti-mutagenicity, and anti-complementary activity as described above. Also, it is necessary to investigate immunopotentiating activity in various aspects, not only the proliferation of immunocompetent cells such as splenocyte, lymph node, or Peyer's patch, but also macrophage activation and cytokine production to describe complex and various immune actions. In particular, Peyer's patches, which are intestinal immunocompetent cells, have been known as induction site for IgA production (Sminia et al., 1983) and thus it is necessary to verify the cell proliferation effect for this part.
Thus, this study was performed to investigate the effects of yam mucilage, ginseng extract (Lee et al., 1993), which has already been known to have immunopotentiation effect, and Bifidobacterium (Lee et al., 1993) in vitro on the proliferation of several types of lymphocytes (splenocyte, lymph node, Peyer's patch) and also to investigate the immunopotentiation effect of yam mucilage by measuring the degree of inducing cytokine (TNF-α & IL-6) production, which is an important mediator in macrophage (RAW 264.7) model system.
Materials and Methods
Preparation of yam mucilage
Short yam used in the experiment was purchased that was cultivated and stored or in the form of dried powder in Bukhu-myeon, Andong city. Short yam was washed and clean yam itself without peeling was used and thinly sliced into 0.5 cm sections to improve the efficiency of extraction, and extracted by hot water extraction at 95-100℃ for 2 hours, and filtered through cotton cloth first to eliminate the remnants and then filtered by vacuum filtration to obtain clean extraction. For dried yam powder, it was hydrated to 10% concentration and then centrifuged to obtain the supernatant mucilage for which hot water extraction was performed. Short yam was made of 82.1% water, 1.52% crude protein, 0.48% crude fat, 0.23% crude fiber, 0.88% crude ash, and the soluble solids was 16.94%. Extracted samples were treated with ammonium sulfate or ethanol to eliminate proteins and went through ultra-filtration or dialysis after being treated with α-amylase, and then passed through ion exchange resin to obtain polysaccharide fraction. The peaks of sugars were measured by phenol-sulfuric acid method for each fraction and identified by PAS-staining. Extracts and mucopolysaccharide fractions were concentrated by a rotary evaporator and then stored at -20℃ for sample analysis and immunostimulating activity measurements.
Preparation of ginseng extract
Ginseng was purchased as 6-year-old fresh ginseng grown in Punggi, Gyeongsangbuk-do and stored at about 0-5℃ and used within 2 months of purchase. Ginseng was washed and then clean ginseng itself without peeling was used. It was sliced into thin sections to improve the efficiency of extraction and extracted by hot water extraction at 95-100℃ for 2 hours, with the ratio of sample : water as 1:40, and filtered through cotton cloth first to eliminate the remnants and then filtered by vacuum filtration to obtain clean extraction. Each extract was concentrated by a rotary evaporator (Eyela) and its solid contents were measured and then stored at -20℃ for use as a sample for animal cell culture.
Bifidobacterium
B. adolescentis M101-4 was purchased from Japan Bifidus Foundation and subculture and main culture were performed in MSR medium (Difco Laboratories) containing 5% lactose at 37℃. After growth curve was drawn, it was cultured until the end of the exponential phase and then centrifuged at 1000×g, 4℃ for 15 minutes to recover cells, and suspended in Hanks' buffered salt solution (Sigma Chemical Co.) to the colony of desirable concentration. The vital count of Bifidobacterium was measured under the anaerobic culture at 37℃ for 48 hours using Gas-Paks (BBL Microbiology Systems). It was heat-treated at 95℃ for 30 minutes and stored at -20℃ for further use as a sample in animal cell culture.
Lymphocyte proliferation
Mice were purchased from Daehan Biolink Co., Ltd. as 8-week-old male BALB/c and acclimated for 1 week after arrival in the animal room and then used in the experiment. 50 µg of bovine serum albumin (Sigma Chemical Co.) emulsified with PBS and complete Freund's adjuvant (Bacto-adjuvant complete H37Ra; Difco Laboratories) was primed subcutaneously on the hind-limb sole and tail of mice, which were then dissected after 2 weeks and lymph nodes were separated. Single cell suspension from separated lymph nodes was made with the RPMI 1640 medium (Sigma Chemical Co) containing 10% (v/v) fetal bovine serum (FBS; Gibco Laboratories) and 50 µM 2-mercaptoethanol and then stained with trypan blue to measure the concentration of viable cells. Lymphocytes of 4×105 cells/well and certain concentrations (1, 5, 25 µg/mL) of samples were placed in a 96-well plate (Costar) and incubated for 72 hours at 37℃, 5% CO2. At this time, for control group, B-cell mitogen lipopolysaccharide (LPS; from Salmonella typhymurium, Sigma Chemical Co.) with 0.1, 1, 10 µg/mL concentrations and T-cell mitogen Concanavalin (ConA; Type IV, Sigma Chemical Co.) with 1, 2, 4 µg/mL concentrations were added. Also, B. adolescentis M101-4 strain, which has been known to have lymphocyte proliferation effect, was added in 106, 107, 108 cells/mL concentrations to compare the degree of cell proliferation stimulation. Cell proliferation was measured by the modification of MTT assay by Mosmann (1983). That is, after lymphocytes and each sample were incubated for 72 hours, 10 µL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT; Sigma Chemical Co.) solution (5 mg/mL PBS) was added and incubated another 6 hours and then 100 µL of isopropanol was added. Absorbance of the solution was measured at 570 nm using a microplate spectrophotometer (Spectra Max 340PC; Molecular Devices).
Splenocyte and Peyer's patch cell proliferation
Spleen and Peyer's patch were separated from unprimed normal mouse (BALB/c, 8-week-old male) and single cell suspension was made in the same manner as for lymphocytes. The separation of Peyer's patch was done by the method of Suzuki et al. (1990). Splenocytes or Peyer's patch cells of 5×105 cells/well concentration and certain concentrations of sample or B. adolescentis M101-4 strain were placed in a 96-well plate and incubated for 48 hours at 37℃, 5% CO2. At this time, for control group, LPS and ConA were used to compare the degree of proliferation stimulation, and cell proliferation was measured by MTT assay.
Macrophage cell line culture & cytokine production
RAW 264.7 cells were purchased from American Type Culture Collection. Cells were subcultured in the RPMI 1640 medium containing 1% (v/v) NTCT 135, 10% FBS, 50 µM 2-mercaptoethanol, 1 mM sodium pyruvate, 100 U/mL penicillin and 100 µg/mL streptomycin, and 5×105 cell/mL of cells and certain concentrations of sample or B. adolescentis M101-4 strain were placed in a 48-well plate and incubated for 48 hours at 37℃, 5% CO2. At this time, samples with or without 1 µg/mL of LPS were compared. After the incubation, the supernatant was collected by centrifugation and used while storing at -20℃ for the quantification of cytokine.
The amount of cytokine in the supernatant was measured by modifying the method of Dong et al. (1994). That is, 1 µg/mL of IL-6 or TNF-α antibodies (rat anti-mouse; PharMingen) diluted with 0.1 M sodium bicarbonate buffer (pH 8.2) was placed in a microtiter plate (Nunc) as 50 µL/well and coated at 4℃ overnight. Plates were washed with 0.01 M PBS (pH 7.2) containing 0.2% (v/v) Tween 20 (PBST) 3 times to eliminate remaining antibodies, and blocked for 30 minutes at 37℃ by adding 300 µL of 3% (w/v) BSA solution, and then washed with PBST 3 times to prevent the nonspecific binding. Next, standard recombinant IL-6 and TNF-α or incubation supernatant were placed as 50 µL/well and reacted for 1 hour at 37℃, and washed again with PBST 4 times, and then reacted for 1 hour at room temperature by adding 50 µL of 2 µg/mL diluted biotinylated rat anti-mouse IL-6 or TNF-α antibody. Plates were washed with PBST 6 times and 1.5 µg/mL streptavidin-horseradish peroxidase conjugate (Sigma Chemical Co.) was added as 50 µL/well and reacted for 1 hour at room temperature, and then washed 8 times, and then the amount of bound peroxidase conjugates was measured by adding 100 µL of substrate solution containing 0.1 M citric-phosphate buffer (pH 5.5) and 0.4 mM of tetramethylbenzidine (Sigma Chemical Co.). Finally, the reaction was stopped by adding the same quantity of 6N-H2SO4 and the absorbance was measured at 450 nm using a microplate spectrophotometer (Spectra Max 340PC; Molecular Devices). The amount of cytokine was calculated from the standard curve using Softmax curve-fitting program (Molecular Devices, Sunnyvale, CA).
Results and Discussion
Lymphocyte proliferation
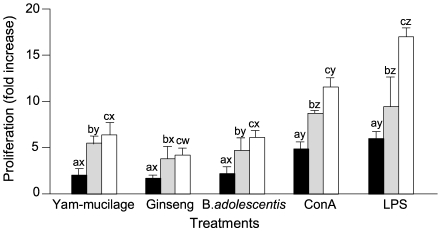
Lymph nodes were separated from BSA primed mice and cell proliferation for each sample was compared. Results were expressed as mean ± SD of data obtained from experiments repeated 3 times, by calculating the degree of proliferation to the control (non-treated group) as fold increase. As shown in Fig. 1, the proliferation effect of yam mucilage fraction was 4.1 fold increase when added in 1 µg/mL compared to control, 6.2 fold increase in 5 µg/mL, and 10.9 fold increase in 25 µg/mL, showing the proliferation increase as the added concentration increased. Not only B-cell mitogen LPS but also T-cell mitogen Con A showed great cell proliferation effect even at low concentrations, which suggests that proliferation occurred not only in B-cells contained in lymph nodes but also in T-cells for primed antigen (BSA). The effect of yam mucilage fraction was weak compared to those of mitogens such as Con A and LPS but higher than that of ginseng extract. For B. adolescentis, it was difficult to compare at the same concentration because heat-treated dead bacterial cells were added, but showed similar proliferation effect as 108 cells/mL Bifidobacteria when added in 25 µg/mL.
Splenocyte and Peyer's patch cell proliferation
Cell proliferation was compared in splenocytes and Peyer's patch cells as same as in lymph node cells. As presented in Fig. 2 and Fig. 3, for splenocytes, the proliferation effect of yam mucilage fraction was 5.0 fold increase when added in 1 µg/mL compared to control, 10.0 fold increase in 5 µg/mL, and 14.7 fold increase in 25 µg/mL, and for Peyer's patch cells, the polysaccharide, promoted the increase when added in 1 µg/mL compared to control, 5.3 fold increase in 5 µg/mL, and 6.4 fold increase in 25 µg/mL. Yam mucilage fraction showed dose-dependent cell proliferation for these lymphocytes, and its effect was weaker than those of mitogens such as Con A and LPS but higher than that of ginseng extract. Also, it showed similar results compared to Bifidobacteria, which has been known to have immunostimulating effect. In particular, because it showed proliferation effect on Peyer's patch cells, an intestinal immunocompetent organ, the effect of yam mucilage fraction as a foodstuff in the intestine might be expected. It has been reported that ginseng extract (Lee et al., 1998), with its components such as saponin and proliferation of not only B-cell or T-cell line but also B or T lymphocytes separated from the spleen or lymph nodes of mice, and it also has been known that lactobacillus such as Bifidobacterium (Lee et al., 1993) has immunoreactivity by cell wall polysaccharides or soluble polysaccharides of the cytoplasm. Also in this study, yam mucilage fraction showed immune cell proliferation effect and its proliferation pattern was similar to that of LPS, and thus it is considered that it might have a mitogen-like function specific to B cells.
Cytokine production by macrophage cells
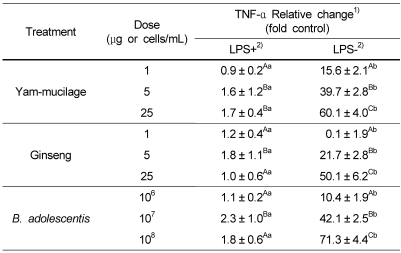
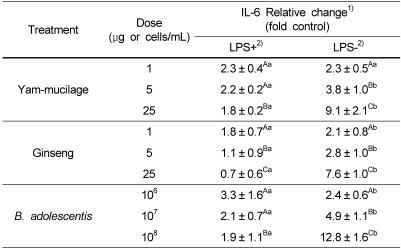
The degree of inducing cytokine production, an important mediator in immune function, in macrophage (RAW 264.7) model by yam mucilage fraction, ginseng extract, and Bifidobacterium was measured by ELISA method. As shown in Table 1, yam mucilage did not promote TNF-α production for macrophage pre-stimulated with 1 µg/mL of LPS, but promoted TNF-α production for LPS-unstimulated macrophage by 15.6 fold when added in 1 µg/mL compared to control, 39.7 fold in 5 µg/mL, and 60.1 fold in 25 µg/mL. Another cytokine, IL-6, produced by macrophage showed relatively weak effect compared to TNF-α, promoting about 9.1 fold increase in IL-6 production when 25 µg/mL was added (Table 2). From the above results, it is thought that yam mucilage fraction directly stimulated macrophage to produce cytokines and thus it is considered that in vivo experiments or the separation of substance should be performed in the future.
Recently, several studies on immunopotentiating activity from plant source were occurred. Lim et al. (2007) reported that water extracts of Acanthopanax divaricatus var. alveofructus and Codonopsis lanceolata showed a significant immunomodulatory activity such as production of IgG antibodies. Kim et al. (1998) reported that Platycodon radix (Changkil) extracts increased natural killer cells in lymphocytic composition against Sarcoma-18O ascites and solid tumor transplanted in ICR mice. Kwon et al. (2001b) reported that MeOH extracts of Schizandra chinensis fructus increased the proliferation of splenocytes and thymocytes. Immunopotentiating substances such as lentinan (Chihara et al., 1987) in mushroom and cell wall components of lactobacillus (Damais et al., 1975; Sekine et al., 1994) have been reported, and thus the relationship between the structure of immunopotentiating substance and its functions need to be investigated for yam mucilage fraction, and the development of functional food or new materials using this is expected for the prevention of decreased immunoactivity due to aging or diseases.


 XML Download
XML Download