

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
HMG coA reductase inhibitor, statin is recognized as one of the most popular drugs for lowering cholesterol in human. Among US adults aged ≧ 40 years with hypertension, 14.5% are taking statin medication in NHANES (1999-02) (King et al., 2007) and an increasing number of the population in Korea are using statin therapy. Besides the cholesterol lowering effect, statins are known to be effective in prevention of cardiovascular diseases such as stroke (Mastuno et al., 2004), myocardial infarction (Susic et al., 2003) and hypertension (Zacco et al., 2003). Beneficial effects of statin further include preservation of bone mineral (Staal et al., 2003), prevention of beta amyloid accumulation (Pentaceska et al., 2002) and cell apoptosis (Kaneta et al., 2002). However, statin associated adverse effects also cannot be ignored. The most serious and frequent side effects from statin use are muscle dysfunctions such as myalgia and cardiomyopathy which may be associated with the function of coenzyme Q10 (CoQ10) in mitochondria electron transport system (ETS). CoQ10 enables the oxidative phosphorylation in the bioenergetic system via its action in mitochondrial ETS of active muscles. Statin therapy has been reported to deplete coenzyme Q10 which shares the cholesterol biosynthetic pathway. Statin therapy caused a reduction of plasma CoQ10 and ubiquinol-10 along with decreased total and LDL cholesterol (Mabuchi et al., 2005). Statin also depleted CoQ10 in platelets and lymphocyte in rats (Littarru et al., 2007) and hepatocyte mitochondrial CoQ10 level in vitro (Tavintharan et al., 2007). Statin user with myopathic symptoms had revealed lower muscular CoQ10 concentration (Lamperti et al., 2005) and supplementation of CoQ10 reverses the decreased plasma CoQ10 and alleviated the myalgic symptom following statin therapy (Nawarskas, 2005). Besides the function in the respiratory chain, the reduced form of CoQ10 (CoQH2), ubiquinol in plasma membrane scavenges radicals generated in metabolic processes, protecting cell membrane from oxidative damage and depletion of coenzyme Q10 leads to a loss of antioxidant capacity. Dhanasekaran et al. (2005) proposed a lipophilic antioxidant, CoQ10 participates regeneration of alpha-tocopherol and ascorbate in the plasma membrane and prevents lipid peroxidation. The extent of oxidative membrane damage has been measured by hemolysis test (Draper et al., 1969) or erythrocyte Na passive leak which can be increased upon exposure to oxygen generating system (Garay et al., 1983). Platelet aggregation and its release reaction were affected by the condition of oxidative stress (Naseem et al., 1999), and platelet superoxide and TXA2 generation were reduced by plant antioxidant (Ryszawa et al., 2006). Green tea polyphenol, eipigallo catechin gallate (EGCG) prevented erythrocyte membrane from lipid peroxidation in vitro (Saffari et al., 2004) and green tea catechins inhibit AAPH induced conjugated diene formation and LDL oxidation in vitro (Osada et al., 2001)
We proposed to test the cholesterol lowering effect of statin in hypercholesterolemic rats and to test whether dietary supplementation of coenzyme Q or green tea can compensate the deplete state of antioxidant following statin therapy, giving better antioxidant index on erythrocyte membrane and platelet aggregation.
Materials and Methods
Animals and diets
Forty of eight week old Sprague Dawley rats (Orient Bio Co Ltd, Gapyung, Korea) were divided into four groups and fed the following diets: 0.5% cholesterol based control diet; control diet plus 30 mg/kg simvastatin; statin plus 15 mg/kg BW CoQ10; statin plus 5% green tea powder (Table 1). Rats had free access to water and were housed in individual cages in a room maintained at 20-25℃ with a 12-hour dark-light cycle. After 4 weeks ad libitum feeding, blood samples were obtained by cardiac puncture into heparinized vacuum tubes, and platelet aggregation and erythrocyte Na efflux were performed with fresh blood. Liver samples were prepared for microscopic examination and plasma and liver samples were stored at -70℃ for later assays.
Plasma and liver lipid assays
Plasma total cholesterol and methanol for triglyceride were assayed using enzymatic kits (Asan Pharmaceuticals, Korea).
Liver lipids were extracted by a modified Folch method (Folch et al., 1957). One gram of liver tissue was homogenized for 5 min in 6 ml of Folch solution [chloroform(2):methanol(1)] and 2 ml H2O. After centrifugation for 10 min, the lower phase which contains liver lipids was separated. Lower phase of lipid fractions after being treated with triton X-100: chloroform (25 µl:475 µl) for total cholesterol and methanol for triglyceride was assayed using enzymatic kits (Asan Pharmaceuticals, Korea).
Platelet aggregation
Platelet aggregation was measured using a Chronolog Whole Blood Aggregometor (model 500-Ca, Havertown, Pennsylvania, USA), of which the instrumental principle is based on the increase in impedance (Ω) across two platinum electrodes as platelet aggregation proceeds. The whole blood was diluted with isotonic saline (1:4) to give platelet concentration of approximately 200,000 platelets/µl. Adenosine diphosphate (ADP, 2 µM) was added to initiate aggregation, and three readings of impedance changes were averaged for each rat.
Erythrocyte Na-leak
Sodium leak is defined as the sodium efflux through passive diffusion with inhibition of ouabain sensitive Na-pump and furosemide sensitive Na-K cotransport and can be increased upon membrane damage of erythrocyte after exposure to free radical generating system such as AAPH [2,2'-azobis(2-Amidino-propane) dihydrochloride]
Blood was centrifuged at 1,000 × g for 10 min, and the plasma and buffy coat were removed. Red blood cells were washed 5 times with a cold isotonic washing solution [150 mM choline chloride, 10 mM Tris-MOPS (morpholinopropane sulfonic acid), pH 7.4 at 4℃], centrifuging at 1,000 × g for 5 min after each wash. The final erythrocyte pellet was resuspended in the choline chloride washing solution to give 40-50% hematocrit. The hematocrit was measured. A 5 µl aliquot of the suspended erythrocyte was added to 5 ml of 0.02% acationox (a metal free detergent, Scientific Products, McGaw Park Illinois, USA) to be used for determination of Na concentrations.
Erythrocyte suspensions of 2 ml each were added to 40 ml Na efflux medium (150 mM choline chloride, 10 mM glucose, 1 mM ouabain, 1 mM furosemide, 10 mM Tris-MOPS pH 7.4 at 37℃) with and without 1 mM AAPH, then mixed gently and aliquoted to 12 tubes. Tubes in duplicate were transferred to an ice bath after incubation in 37℃ shaking water bath for 0, 10, 20, 30, 40 and 50 min. Tubes were centrifuged at 1,000 × g for 5 min, then the supernatants were removed and measured Na concentration. Na leak (mmole/l rbc/hour) is calculated from efflux slope which increases upon exposure to free radicals.
Calculations (Kang et al., 1990)
Na µg/(ml×min)×60 min×mmole/23 mg×[44 ml-(4 ml×hct)/(0.0041×hct)]×(mg/1000 µg) = mmole/l rbc/hour (Na leak)
Na µg/(ml×min)×mmole/23 mg×101/hct×(mg/1000 µg) = mmole/l rbc (Intracellular Na)
Hemolysis
Autohemolysis was performed by modified method of Draper et al. (1969); determining the extent of hemolyis with the absorbance at 415 nm using a spectrophotometer.
Liver and PRP TBARS production
Platelet rich plasmas (PRP) were obtained after centrifuging whole blood at 300 × g for 10 minutes. PRP and liver samples were preincubated at room temperature for 48 hours before determining TBARS. Thiobarbituric acid reactive substance (TBARS) was determined according to a modified Yagi's method (Yagi, 1976). Plasma lipids were precipitated with phosphotungstic acid and TBARS in the pellet were measured at excitation 515 nm, emission 553 nm with a fluometer (Kontron model SFM 25) using 1,1,3,3 tetraethoxypropane (Sigma-Aldrich) as a standard.
Histological examination
Two parts of liver samples each rat were obtained from 4 rats each group. After being washed with saline, samples were fixed in 10% formalin for 48 hours and dehydrated in xylene for 18 hours. Samples were then embedded in paraffin and cut into 4-5 um cross section. Tissues were stained with hematoxylin-eosin for microscopic observation.
Results
Weight gain and food efficiency
Weight gain and food efficiency are shown in Table 2. The final body weight was decreased in the statin group compared with the control and increased in groups with CoQ10 and green tea to the level of the control, but no statistically difference. Food intake was not different among groups, but FER decreased in statin group and increased in green tea group, showing statistical difference between these two groups (p<0.05).
Plasma and liver cholesterol and triglyceride
Plasma total cholesterol (Table 3) were somewhat increased in the statin group compared with the control, and CoQ10 or green tea further increased these levels, but was no statistical difference among groups. HDL-cholesterol was decreased in the simvastatin group and increased in groups with CoQ10 and green tea. Plasma triglyceride was decreased in the statin group and increased in groups of CoQ10 and green tea, and the difference between groups of statin and green tea was significant (p<0.05). Liver total cholesterol (Table 3) was not different between the control and statin group, but was significantly decreased in the group with green tea compared with other groups (p<0.05). Liver triglyceride was decreased in groups of statin and green tea compared with the control, and the difference between groups of the control and green tea was significant (p<0.05). The simvastatin treatment somewhat decreased plasma and liver triglyceride without affecting plasma or liver cholesterol. Green tea increased HDL-cholesterol and decreased the liver levels of total cholesterol and triglyceride.
Whole blood platelet aggregation
Platelet aggregation (Table 4) of both the initial slope and the maximum was not statistically different among groups, but the group with green tea was tended to be higher in initial slope and lower in the maximum compared with other groups.
Hemolysis and erythrocyte Na leak
Hemolysis and erythrocyte Na leak are shown in Table 5. Hemoysis was increased in all groups with simvastatin compared to the control, but was not statistically significant. Intracellular Na of group with green tea was significantly higher than control or statin group (p<0.05). Na leak in intact cells was significantly decreased in the statin group compared with control (p<0.05). Na leak in AAPH treated cells was also significantly reduced in the statin group compared with the control and group with CoQ10 (p<0.05). Increase in Na leak after treating AAPH was the highest in the group with CoQ10. Na leak in intact cell was not correlated with intracellular Na.
Liver and PRP TBARS production
Liver TBARS production (Table 6) was increased in the statin group and decreased in the group with green tea, but no statistical difference among groups. TBARS production in platelet rich plasma was significantly decreased in groups with CoQ10 and green tea compared with the control and statin groups (p<0.05).
Discussion
Akiba et al. (1999) observed that NK-104, a HMG CoA reductase inhibitor suppressed body weight gain or decreased body weight with dose of 50 mg/kg in rats. Chronic pravastatin plus captopril treatment decreased weight gain, improving lipid profile and insulin resistance (Villa et al., 1998). Farmer et al. (2007) reported that a significant reduction of weight and total-cholesterol was observed in rosuvastatin-treated wild type mice with the reduced number of cells in the hippocampus labeled with glial fibrillary acidic protein (GFAP) where rosuvastatin may function in decreasing appetite. In the present study, the group with simvastatin decreased weight gain without reducing food intake. CoQ10 increased food intake, while green tea increased food efficiency ratio, both increasing weight gain in statin treated rats. Unlike green tea extract which showed antiobese effect in our preliminary experiment, green tea powder gave a normal weight gain in this experiment, which suggest green tea cellulose may help with weight gain increasing FER.
Hypochoelsterolmic effects of statin have been reported in human and animal studies. Fluvastatin, 80 mg daily for 8 weeks reduced LDL cholesterol by 35% in patients with type II hypercholesterolemia (Scharnagl et al., 2006). Saito et al. (2003) reported rosuvastatin with daily dose rage of 1-40 mg showed a linear response in reduction of total cholesterol and apolipoprotein B in hypercholesterolemic patients. Fluvastatin, 3.5 mg/kg BW decreased plasma total cholesterol and triglyceride in mice (Galus et al., 2006) and lovastatin, 5 mg/kg BW or pravastatin 2.5 mg/kg BW reduced plasma cholesterol by 36%, 30% respectively with reduction of HMG CoA reducase by 50% and 41% in chow fed dog (Davis et al., 2001). Fluvastatin of 2 mg/kg BW daily decreased plasma total cholesterol and triglyceride with an increased HDL-cholesterol in rabbits (Bandoh et al., 2000). Aggawal et al.(2005) reported that atorvastatin at 50mg/100g diet in guinea pigs decreased 35% LDL cholesterol and 40% triglyceride with decreased cholesterol ester transfer protein (CETP) activity. Unlike these results, Sawada et al. (2002) reported that pravastatin (1~100 mg/kg BW) in hamsters increased serum cholesterol levels and dose dependently decreased serum triglyceride levels, and pravastatin at 3.2 and 10 mg/kg increased hepatic HMG-CoA reductase activity. The species difference in the metabolic pathway of cholesterol and lipoprotein can explain for this discrepancy. Approximately 65% of total cholesterol in human is endogenously synthesized in extrahepatic tissues (Dieschy et al., 1993). Fernandez et al. (2004) observed that like humans, guinea pigs have moderate rates of hepatic cholesterol synthesis, whereas the major site of cholesterol synthesis in rats is the liver, and guinea pigs, unlike rats, carries most of cholesterol in LDL and upregulation of LDL receptor by statin drug is another way in lowering cholesterol in humans and guinea pigs. Intestinal and hepatic acyl CoA:cholesterol acyl transferase (ACAT) which are not active in rats were inhibited by statin in guinea pigs resulting in significant reduction of plasma cholesterol (Fernandez et al., 2001). Intestinal and hepatic ACAT are known to be involved in cholesterol absorption, storage and lipoprotein assembly, producing cholesterol ester depleted chylomicron and VLDL particles and reducing plasma cholesterol. In present study, simvastatin with high dose of 30 mg/kg BW failed to reduce plasma and liver total cholesterol with minor reduction of plasma and liver triglyceride. Many of enzymes or proteins such as HMG CoA reductase, CETP, ACAT, cholesterol 7α hydroxylase and more recently apical sodium dependent bile acid transporter (ASBT), microsomal transport protein (MTP) are known to be involved in metabolic pathway of lipoprotein, and more studies at molecular level are needed to clarify the metabolic differences between species. Mabuchi et al. (2007) reported CoQ10 supplementation of 100 mg/day did not affect the plasma level of LDL cholesterol and triglyceride, but significantly increased HDL cholesterol in hypercholesterolemic patients with atorvastatin therapy, which agreed with our results in some extent. We assume that supplemented CoQ10 in our study did not exert feedback inhibition on HMG CoA reductase in cholesterol synthesis. Raederstorff et al. (2003) reported tea catechin prevents lipid absorption, decreasing plasma total cholesterol and triglyceride. Green tea powder of 4% in rat had antiobese and hypotriglycemic effect without influencing plasma cholesterol (Sayama et al., 2003). Dietary green tea in the present study significantly decreased the liver levels of cholesterol and triglyceride, and the increased plasma triglyceride in the group of green tea can be explained with the increased secretion of liver triglyceride, preventing fatty liver. Liu et al. (1995) observed that increased liver triglyceride with decreased plasma triglyceride in hypercholesterolemic rat was attributed to the low secretion of de novo synthesized triglyceride, causing fat accumulation in the liver.
Platelet aggregation and its release reaction were affected by the condition of oxidative stress (Naseem et al., 1999). The release reaction in the process of aggregation induces the 2nd phase of aggregation where the released ADP and serotonin potentiate platelet aggregation. Pravastatin treatment significantly reduced LDL-cholesterol and inhibited ADP induced platelet aggregation and TXB2 synthesis in Chinese patients with primary hypercholesterolemia (Dajani et al., 2002). Haramaki et al. (2007) reported that in vitro study fluvastatin dose-dependently inhibited platelet aggregation and decreased platelet-derived nitric oxide (PDNO) release. Statin may function on platelet aggregation through its action on cholesterol metabolism and membrane lipids, since hypersensitivity of platelet is associated with hypercholesterolemic condition. CoQ10 in form of ubiquinol may reduce oxidative stress which leads to hypersensitivity of platelets. Green tea suppressed the collagen induced platelet aggregation independent manner of cholesterol lowering. In present study, platelet aggregation in initial slope and maximum aggregation was not affected with statin or CoQ10, but green tea tended to decrease the initial slope and increase the maximum aggregation which is associated with the platelet survival after aggregation.
Atorvastatin decreased cholesterol contents in erythrocyte membrane decreasing cholesterol/phospholipid ratio, giving membrane stability with increased Na-K ATPase in guinea pig (Uyuklu et al., 2007). Cholesterol content of membrane is one of determinant factors for membrane fluidity. Higher concentrations of membrane cholesterol and TBARS and lower activity of Na-K ATPase were observed in erythrocyte of patient with hyperlipidemia, and pravastatin reversed or lessened this condition independent manner of cholesterol lowering effects (Broncel et al., 2007). In the present study, simvastatin, a lipophilic statin, can cross membrane layers and act on controlling membrane cholesterol and membrane stability resulting in decreased passive diffusion in intact cells. CoQ10 and green tea somewhat increased the reduced passive Na leak in intact cells of the statin group. The magnitude of Na leak of intact cells is theoretically correlated with intracellular Na, but the control group had the highest passive Na leak with the lowest intracellular Na concentration. Leak increase (Δ Na leak) after treated with AAPH was the highest in the group with CoQ10, which can be interpreted as the more membrane damage. CoQ10 may not be effective in stabilizing membranes. Broncel et al. (2006) suggested statin induced antioxidant enzymes such as CAT, SOD and glutathione peroxidase, reducing plasma lipids and TBARS production. Atorvastatin reactive metabolite (ATM) blocks membrane cholesterol from oxidative stress decreasing lipid peroxidation in LDL (Mason et al., 2006). Plant antioxidant reduced platelet superoxide and TXA2 production (Ryszawa et al., 2006). CoQ10 and green tea decreased TBARS production in liver and PRP, but CoQ10 was not effective in protecting membrane from oxidative stress. Erythrocyte Na leak is Na efflux through lipid bilayer of cell membrane in the manner of passive diffusion by concentration gradient. Upon exposure to AAPH radical generating system, Na leak would increase with the extent of membrane damage. TBARS is a product of membrane lipid peroxidation, and TBARS production will be correlated with leak increase. The leak difference between intact cells and AAPH treated cells was the highest in rats with CoQ10, coenzyme Q10 in the present study did not prevent membrane damage, but it was effective in reducing TBARS production in PRP. Antioxidant effect of green tea was obvious, but coenzyme Q10 did not show consistent effects. More studies are needed to elucidate the mechanism of CoQ10 functioning as antioxidant.
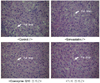
We examined the liver tissue for some damages from the high dose of statin treatment. Since all groups of experimental animals received a cholesterol diet, there was no difference among groups on gross observation. Livers from all groups were large with LW/BW ratio raging 4.2-4.3 compared with the rough value of 2.5 in normal rats and the liver tissue was quite hypochromic, which can be defined as fatty liver. On microscopic observation (Figure 1), liver specimens from groups of CoQ10 and green tea showed less fat drops than those from the control and statin group, which suggests that CoQ10 and green tea alleviate fatty liver in some extent in hypercholesterolemic rats.
In the present study, simvastatin treatment in hypercholesterolemic rats did not show a cholesterol lowering effect, but showed a hypolipidemic effect. CoQ10 supplementation to the statin diet did not affect plasma and liver levels of cholesterol and triglyceride. Green tea supplementation to the statin diet prevented the liver fat accumulation caused by high cholesterol diet. CoQ10 and green tea did not affect platelet aggregation and membrane Na leak, but reduced TBARS production in liver and platelet rich plasma.
The beneficial effect of CoQ10 and green tea as an antioxidant may not be further discussed unless CoQ10 depletion with statin treatment is verified in rat model. Numerous clinical trials have been done on CoQ10 supplementation at dose of 90 mg-600 mg/day depending on the clinical purpose in patients with statin therapy, mostly showing beneficial effects of statin. Studies reported that CoQ10 supplementation recovers the depleted plasma CoQ10 following statin therapy and alleviated myopathic and myalgic symptoms. The council of responsible nutrition, USA reported that a daily dose of 1200 mg/day CoQ10 was tolerated in most patients with statin therapy, suggesting the observed safety level (OSL) upper 1200 mg/day. However, as long as there are reports of nausea and other adverse gastrointestinal effects in individuals taking CoQ10 at the low level as 60 mg-200 mg/day, extra CoQ10 supplementation may not be yet recommendable to the general population without statin therapy.






 XML Download
XML Download