

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Circulating leukocytes adhere to endothelial cells and undergo transmigration through the vessel wall into the extracellular matrix. Leukocyte extravasation requires the secretion of matrix-degrading enzymes, matrix metalloproteinases (MMP), which allow leukocytes to penetrate through the basement membrane. MMP can be divided into four categories based on substrate preference: collagenases, gelatinases, stromelysins and membrane-associated MMP. MMP-2 and MMP-9 preferentially degrade denatured gelatin and native collagen type IV which are one among the main components of basal membranes. Transmigration and metastasis of macrophages and tumor cells require disruption of basement membrane, so that MMP-2 and MMP-9 have been suggested to play a critical role in such processes (Galis et al., 1994; Nagaoka & Hirota, 2000; Saarialho-Kere et al., 1993).
The vascular endothelial cells are an important target of pro-inflammatory cytokine TNF-α (Pober & Cotran, 1990; Ross, 1993), in which it modulates endothelial expression of many genes involved in cell adhesion, thrombosis and inflammatory responses leading to atherosclerosis (Krasinski et al., 2000). Adhesion molecules such as vascular cell adhesion molecule-1 (VCAM-1) and intercellular cell adhesion molecule-1 (ICAM-1) are shown to play an important role in the induction of inflammation (Berk et al., 2001; Nakashima et al., 1998). The VCAM-1 induction of is a common feature in inflammatory environments and occurs in the early development of atherosclerosis (Schonbeck et al., 2000).
Several epidemiological studies have shown that red wine consumption is inversely related to cardiovascular disease (Aviram M & Fuhrman B, 2002; Belleville, 2002). Subsequently, multiple experiments have been conducted to demonstrate the pathophysiological activities of various components present in red wine. Resveratrol, a polyphenol subsisting in multiple plants including grapes and mulberries, is thought to be as a potential protective component (Bertelli et al., 1995; Hao & He, 2004). There is growing evidence that resveratrol can prevent or delay the onset of various cancers, heart diseases, ischemic and chemically induced injuries, pathological inflammation and viral infections (Baur & Sinclair, 2006; Bradamante et al., 2004; Shankar et al., 2007). In a recent report (de Lange et al., 2007), red wine inhibited platelet functions through content of polyphenolic grape extract that stimulated the inhibitory receptor, platelet endothelial cell adhesion molecule-1 (PECAM-1), thereby attenuating platelet activation. In addition, resveratrol suppressed monocyte chemotactic protein-1 binding activity of THP-1 cells and its receptor CCR2 mRNA, indicating that these inhibitory effects of resveratrol on chemokine receptor binding and expression may contribute, in part, to its cardiovascular protective activity in vivo (Cullen et al., 2007).
On the basis of the literature evidence that resveratrol is a protective component preventing the onset of pathological inflammation, the present study assessed the anti-inflammatory activity of resveratrol with respect to VCAM-1 expression in TNF-α-exposed human umbilical vein endothelial cells (HUVEC). Furthermore, the leukocyte extravasation was investigated by measuring gelatinolytic activity of MMP and monocyte migration.
Materials and Methods
Materials
M199 medium chemicals, RPMI 1640 medium chemicals, Resveratrol and 3-(4, 5-dimetylthiazol-yl)-diphenyl tetrazolium bromide (MTT) were obtained from Sigma-Aldrich Co. (St. Louis, MO), as were all other reagents, unless specifically stated elsewhere. Collagenase was purchased from Worthington Biochemicals (Lakewood, NJ). Fetal bovine serum (FBS), penicillin-streptomycin, trypsin-EDTA, bovine brain extract, human epidermal growth factor and hydrocortisone were purchased from Cambrex Corporation (East Rutherford, NJ). Antibodies against human VCAM-1, human β-actin and human VCAM-1 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Horseradish peroxidase-conjugated goat anti-rabbit IgG was provided as a secondary antibody by the Jackson ImmunoResearch Laboratory (West Grove, PA). Human monocytic leukemic cell line THP-1 was obtained from American Type Culture Collection (Rockville, MD). TNF-α was obtained from Roche Molecular Biochemicals (Mannheim, Germany).
Preparation and culture of HUVEC
HUVEC were isolated from umbilical cords using collagenase as described elsewhere (Choi et al., 2005). Cultures were maintained at 37℃ humidified atmospheres of 5% CO2 in air. Cells were cultured in 25 mM HEPES-buffered M199 containing 10% FBS, 2 mM glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin supplemented with 0.75 mg/mL human epidermal growth factor and 0.075 mg/mL hydrocortisone. Cells were passaged at confluence and used within 10 passages. Endothelial cells were confirmed by their cobblestone morphology and uptake of fluorescent 1.1'-dioctadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate-labeled acetylated LDL.
HUVEC were plated at 90-95% confluence in all experiments. In experiments for resveratrol dose responses to TNF-α-induced VCAM-1 expression and MMP-2 activity, cells were incubated with 1-50 µM resveratrol overnight and subsequently exposed to 10 ng/mL TNF-α for 6 h. It has been shown by using high-performance liquid chromatography that concentrations of trans-resveratrol in human plasma ranged 15.0-4,000 ng/mL with intra-assay variability of 1.9-3.7% and inter-assay variability of 2.5-4.0% (Zhu et al., 1999).
Culture of THP-1 cells and in vitro cell adhesion assay
HUVEC were grown in 25 mM HEPES-buffered at a density of 1.0 × 105 cells on 4-well glass chamber slides. Human monocytic THP-1 cells were labeled with 5 µM calcein-AM (Molecular Probes Inc., Eugene, OR) in RPMI 1640 medium containing 10% FBS. Cells were pre-treated with 10-50 µM resveratrol overnight prior to the 6 h exposure to 10ng/mL TNF-α. In the co-culture models, the labeled THP-1 cells were seeded at a density of 5.0 × 105 cells onto near-confluent monolayer of HUVEC treated with the resveratrol and/or TNF-α and were incubated for 2 h. After the co-cultured cells were thoroughly washed, photograph images were obtained at 485 nm excitation and 538 nm emission using a SPOT II digital camera-attached fluorescence microscope with Spot II data acquisition software (Diagnostic Instrument, Livingston, Scotland).
Western blot analysis
For the VCAM-1 induction by TNF-α, whole cell extracts were prepared from resveratrol-treated and TNF-α-exposed HUVEC in 1 M Tris-HCl (pH 6.8) lysis buffer containing 10% SDS, 1% β-glycerophosphate, 0.1 M Na3VO4, 0.5 M NaF and protease inhibitor cocktail. Cell lysates with equal amounts of total protein were fractionated by electrophoresis on 8% SDS-PAGE gels and transferred onto a nitrocellulose membrane. Nonspecific binding was blocked by soaking the membrane in TBS-T buffer [0.5 M Tris-HCl (pH 7.5), 1.5 M NaCl, and 0.1% Tween 20] containing 5% nonfat dry milk for 3 h. The membrane was incubated for 3 h with a primary polyclonal rabbit antihuman VCAM-1 antibody after three washes with TBS-T buffer, the membrane was then incubated for 1 h with horseradish peroxidase-conjugated goat anti-rabbit IgG. The protein level of VCAM-1 was determined by using Supersignal West Pico chemiluminescence detection reagents (Pierce Biotech. Inc., Rockford, IL) and Konica X-ray film (Konica Co., Tokyo, Japan). Incubation with monoclonal mouse β-actin antibody was also performed for the comparative control.
Gelatin zymography
Culture medium of HUVEC or THP-1 cells was mixed in the ratio of 5 to1 with sample buffer and electrophoresed in 10% SDS-PAGE containing 1 mg/mL gelatin. Gels were renaturated by exchanging SDS for 2.5% Triton X-100, followed by 20 h incubation at 37℃ in the activation buffer (50 mM Tris, pH 7.5, 10 mM CaCl2, 0.2 M NaCl and 0.05% Brij). Gels were subsequently stained with Coomassie blue staining solution (0.1% Coomassie R250; 45% MeOH; 2% acetic acid) for 1 h, followed by being destained with 30% MeOH and 10% acetic acid.
THP-1 cell migration
The experimental models for the monocyte migration employed 24-transwell inserts with pore sizes of 3 µm (Corning Incorp., Corning, NY). The lower surface of the insert filter was coated with 10 µL of 1 mg/mL gelatin solution and dried for 1 h. THP-1 cell suspensions of 106 cells/mL in serum-free RPMI 1640 were loaded in the upper compartment of the transwell chamber. The cells were treated with 25 µM resveratrol and exposed to 10 ng/mL TNF-α for 48 h. Transmigration was stopped by removing the transwell insert. The number of cells that had migrated to the lower compartment across the lower gelatin surface of the filter was counted microscopically. In addition, photography was obtained using a Motic AE30/31 inverted microscope (Motic Co., Xiamen, China).
Data analysis
The results are presented as mean ± SEM for each treatment group in each experiment. Statistical analyses were conducted using Statistical Analysis Systems statistical software package (SAS Institute Inc., Cary, NC). Significance was determined by one-way ANOVA followed by Duncan multiple range test for multiple comparisons. P values <0.05 were considered statistically significant.
Result
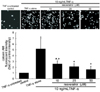
Inhibition of TNF-α-induced monocyte adhesion by resveratrol
The in vitro data for the THP-1 cell adhesion to HUVEC was shown in Fig. 1. A small number of monocytes were adhered to quiescent HUVEC free of 10 ng/mL TNF-α. There was heavy staining in TNF-α-alone-exposed HUVEC, indicative of a substantial increase in the THP-1 cell adherence to the activated HUVEC. However, adding resveratrol at non-toxic doses of 10-50 µM to TNF-α-exposed cells markedly attenuated the monocyte adherence in a dose-dependent manner (Fig. 1). It should be noted that ≥10 µM resveratrol was effective in inhibiting leukocyte adhesion to cytokine-activated endothelium.
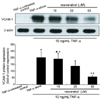
Mitigation of TNF-α-induced expression of VCAM-1 by resveratrol
Western blot analysis was performed to address that resveratrol blocked expression of VCAM-1 induced by TNF-α. Expression of VCAM-1 protein was greatly elevated in TNF-α-stimulated HUVEC over the quiescent cells (Fig. 2). When resveratrol was supplemented in the range between 10 and 50 µM, the enhanced expression of VCAM-1 dose-dependently declined. This data showed that ≥25 µM resveratrol diminished the TNF-α-stimulated expression of VCAM-1. These results imply that other adhesion molecules such as ICAM-1 and PECAM-1 may be involved in blocking the leukocyte trafficking.
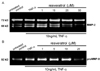
Gelatinolytic activity of MMP-2 and MMP-9
When HUVEC were treated with 10 ng/mL TNF-α under serum-free conditions, the gelatin zymography revealed that the gelatinolytic activity of MMP-2 active form secreted in culture media within 24 h was substantially increased (Fig. 3A). In contrast, the treatment of TNF-α-exposed HUVEC with resveratrol mitigated the increased gelatinolytic activity of MMP-2 active form (69 and 68 kDa) only at its concentrations of ≥25 µM (Fig. 3A). It should be noted that the pro-form of MMP-2 (72 kDa) secretion was not influenced in presence of TNF-α and/or resveratrol. In addition, the enzyme activity of pro-form MMP-9 (92 kDa) secreted from TNF-α-added THP-1 cells was elevated, whereas 25 µM resveratrol markedly suppressed the MMP-9 activity (Fig. 3B). The active form of MMP-9 (82 kDa) was not apparent in TNF-α-exposed HUVEC.
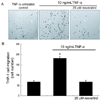
Blockade of TNF-α-induced THP-1 migration by resveratrol.
Fig. 4 showed inhibitory effects of resveratrol on THP-1 transmigration into the vascular matrix tissue. The THP-1 cell migration across the gelatin layer mimicking basement membranes was markedly augmented within 48 h after an addition of TNF-α. On the contrary, the elevated monocyte transmigration was abolished by 25 µM resveratrol, indicating that resveratrol blunted cytokine-stimulated monocyte extravasation effectively through suppressing secretion of gelatinolytic MMP and subsequent degradation of vascular matrix.
Discussion
This study attempted to test the hypothesis that resveratrol, a polyphenol found in red wine and peanuts, inhibits mononuclear leukocyte recruitment on the TNF-α-induced vascular endothelium. Four major findings were observed from this study. 1) Resveratrol at the non-toxic dose of ≥10 µM, nearly abolished THP-1 adhesion onto TNF-α-exposed cells. 2) Adding resveratrol in the dose range between 10 and 50 µM dose-dependently attenuated expression of VCAM-1 up-regulated by TNF-α. 3) When resveratrol at the doses of ≥25 µM was supplemented to TNF-α-exposed endothelial cells, the elevated gelatinolytic activity of MMP-2 was mitigated. In addition, the enhanced activity of MMP-9 was reduced in 25 µM resveratrol-treated THP-1 cells. 4) Resveratrol at the dose of 25 µM suppressed the transmigration of THP-1 monocytes exposed to TNF-α. These overall observations demonstrate that resveratrol has the potential capability to prevent the early events in atherosclerosis. The ability to block the TNF-α-induced monocyte trafficking and extravasation argues for the major target of action of resveratrol.
Numerous studies have focused on beneficial effects of polyphenols on cardiovascular functions including improvement of endothelial function, inhibition of angiogenesis and cell migration and proliferation in blood vessels (Choi et al., 2005; Katiyar, 2006; Middleton et al., 2000; Nijveldt et al., 2001). There is compelling evidence that the distinct structures of polyphenols appear to be partially responsible for their protective functions (Fukuhara et al., 2006; Hu et al., 2006; Teixeira et al., 2005). The mechanisms triggered by polyphenols with specific structures contribute to the vasoprotective, anti-angiogenic, anti-atherogenic, vasorelaxant and anti-hypertensive effects found in animals and in patients (Stoclet et al., 2004). In addition, the current evidence suggests that there is a plausible target for inhibiting atherosclerosis by grapes and/or its polyphenolic constituents through modulating novel pathways involved in the leukocyte extravasation.
Moderate consumption of red wine has been associated with beneficial effects on cardiovascular diseases, which is the phenomenon named as the French paradox (Belleville, 2002; Bertelli et al., 1995; Hao & He, 2004). Resveratrol has been regarded as the potential protective polyphenol present in red wine. In the present study resveratrol blunted monocyte adhesion onto the activated endothelial cells and its transmigration across a gelatin matrix barrier, indicating that resveratrol is capable to block the early process of atherosclerosis. It was reported that resveratrol might improve obesity-induced atherosclerosis by modulating the TNF-α-induced secretion changes of adipokine and adiponectin (Ahn et al., 2007). Additionally, resveratrol suppressed monocyte chemotactic protein-1 receptor CCR2 binding and expression of THP-1 cells responsible for atherosclerosis (Cullen et al., 2007). However, this study did not elucidate possible mechanisms by which resveratrol conferred the beneficial effects in endothelial responses to cytokines during inflammation. It was suggested that resveratrol be an antioxidant protecting endothelial cells from oxidized LDL-induced oxidative stress via direct ROS scavenging and inhibition of NADPH oxidase activity (Chow et al., 2007). In addition, this polyphenol inhibited IL-6-induced ICAM-1 gene expression, in part, by interfering with Rac-mediated pathways via the attenuation of STAT3 phosphorylation (Wung et al., 2005).
MMP-2 and MMP-9 preferentially degrade denatured gelatin and native collagen type IV, which are the main components of basal membranes. Accordingly, transmigration of macrophages requires disruption of basement membrane, and thus MMP-2 and MMP-9 have been implied to play a critical role in such processes (Galis et al., 1994; Nagaoka & Hirota, 2000; Saarialho-Kere et al., 1993). The present study showed that resveratrol interfered with monocyte transmigration concomitantly with dampening MMP secretion of endothelial cells and monocyes. A dose-related decrease of MMP-2 production after resveratrol treatment of human cultured glioblastoma cells revealed that resveratrol may influence the ECM remodeling occurring with tumor invasion (Gagliano et al., 2005). It was also shown that resveratrol inhibited phorbol myristate acetate-induced MMP-9 expression by inhibiting signal transduction of c-Jun N-terminal kinase and protein kinase Cδ (Woo et al., 2004). Considerable interest is increasing with regard to the use of dietary botanical supplements for chemopreventive and neuro- and cardiovascular-protective effects. Indeed, dietary botanicals such as green tea polyphenols and grape seed proanthocyanidins have been considered as MMP inhibitors and as cancer chemo-therapeutic agents (Katiyar, 2006).
In summary, the current study has demonstrated that resveratrol is capable of preventing early processes of atherosclerosis involving inducible VCAM-1 expression and MMP secretion. Resveratrol blocked monocyte adhesion onto the TNF-α-activated endothelium most likely via the down-regulation of VCAM-1 expression. In addition, this compound inhibited matrix-degrading MMP secretion from cytokine-activated endothelial cells and monocytes. This observation might have clinical implications for therapeutic strategies preventing and attenuating inflammatory diseases. Although definite mechanisms underlying the protection of resveratrol against early atherogenic process are not fully defined in this study, the VCAM-1 expression and MMP secretion may be a pivotal target of anti-atherogenic actions of resveratrol.
 XML Download
XML Download