

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
When a large volume of water is ingested and the concentration of sodium (Na+) in arterial plasma (PNa) declines appreciably, water will be excreted promptly1). The signal is swelling of cells in the osmostat, which inhibits the release of vasopressin and thereby, leads to the removal of aquaporin 2 water channels (AQP2) from the luminal membranes of the late distal nephron2). In this setting, the distal delivery of filtrate sets the upper limit on the magnitude of the water diuresis. In quantitative terms, our best estimate of the distal delivery of filtrate is 27 L/day3) and the maximum water diuresis is ~22 L/day4). Hence there is another important factor that contributes to the magnitude of a water diuresis. This study was designed to gain insights into this missing component of a water diuresis.
Results to be reported indicate that close to half of the water reaching the distal nephron segments during water diuresis is reabsorbed. This reabsorption is likely to occur in the inner medullary collecting duct, as permeability to water when vasopressin does not act has been demonstrated in this nephron segment in rats5). We shall speculate that the function of this reabsorption of water is to desalinate the final urine. These principles may provide useful insights into the pathophysiology of hyponatremia in certain patients with syndrome of inappropriate antidiuresis (SIAD)6, 7).
Go to : 
Methods
1. Animals
Adult male Wister rats (weight 300-400 g) were cared for in accordance with the principles and guidelines of the Canadian Council on Animal Care. The Animal Care Committee of St. Michael's Hospital approved the study protocols.
2. Evaluation of water reabsorption downstream from the early distal convoluted tubule during water diuresis
Our objective was to induce water diuresis while minimizing the non-osmotic stimuli for the release of vasopressin. Thus the animals were awake and housed in individual metabolic cages for collection of urine. The drinking solution contained 5% sucrose to encourage the ingestion of a large volume of this solution within 2 hours. Glucose was not detected in the urine in any of these experiments.
Since rats fed their usual chow did not have a UOsm that was less than their POsm after drinking this water load, we reasoned that the effective osmolality of this ingested solution may have risen owing to the addition of electrolytes from the ingested food while in the stomach. Therefore, the studies were repeated after withholding rat chow on the night before the experiment (n=6 rats). Using this modified protocol, all rats had a water diuresis. Once the rate of excretion of creatinine was constant, the composition of the urine did not reflect mixing of fresh urine with more concentrated prior urine in the bladder. We used the ratio of creatinine concentrations in the urine and in plasma ((U/P)Creatinine), which is akin to the (TF/P)Inulin during micropuncture experiments to reflect the volume of filtrate that was reabsorbed in segments of the nephron prior to the site of micropuncture.
3. Measurement of the osmotic driving force to reabsorb water during a water diuresis
The objective was to measure the osmotic driving force across the inner medullary collecting duct by measuring simultaneous UOsm and papillary interstitial osmolality when the UOsm was appreciably lower than the POsm. Rats were placed in metabolic cages for this 2-day protocol. As in the first protocol, food was withheld overnight on day 1. The drinking solution was 5% sucrose to encourage prompt and complete consumption of this fluid. Each rat (n=7) was given 30 mL of this solution at 16:45 hour and again at 22:45 hour on day 1. On the morning of day 2, the rats were given 15 mL of 5% sucrose at 08:45 hour and again at 09:45 hour. The entire volume was ingested in <60 minutes and a brisk increase in urine output was noted. During the period when the urine flow rate was very large, the voided urine was removed at 10:10 hour and at 10:45 hour. Blood and urine were analyzed as described above. A second group of 14 rats was treated in an identical fashion; these rats were sacrificed by decapitation to obtain the renal papilla for analysis at 10:10 hour (n=7) and at 10:45 hour (n=7).
Composition of the renal papilla: The papilla of one kidney was used to measure its content of electrolytes, ammonium (NH4+) and urea, while the papilla from the other kidney was used to measure its content of water by desiccation8). Each kidney was sliced obliquely along its longitudinal axis with a sharp knife to expose the intact papilla. The papillary tip was blotted, excised, and transferred immediately to a pre-weighed plastic vial and sealed. After the vial plus the papilla was weighed, 1 mL of the solution used for the measurement of Na+ and potassium (K+) by flame photometry was added and the tissue was homogenized. This fluid was analyzed for Na+, K+, NH4+, and urea. The osmolality in the excised papilla was calculated after adding the sum of the contents of all the measured osmoles (2×(Na++K++NH4+)+urea) and dividing this value by the content of water per g.
4. Analytical techniques
Na+ and K+ in plasma and urine were determined by flame photometry (Radiometer, FLM-3, London Scientific Ltd., London, ON, Canada), Cl- was determined by electromimetic titration (Chloride meter, CMT 10, London Scientific Ltd., London, ON, Canada), osmolality was measured by freezing point depression (Advanced Instruments Inc, Needham Heights, MA, USA), and gas analysis in blood and urine was performed at 37℃ with a digital pH/blood gas analyzer (Corning 178 blood pH analyzer). NH4+, urea and creatinine were measured as previously described9, 10).
5. Statistical analysis
Results are reported as mean±SEM. Statistical analysis was performed by paired analyses for urine and papillary osmolalities using a paired Student t-test. A P value that was less than 0.05 was considered to be statistically significant.
Go to :
Results
It is essential to understand the following rationale to interpret the results in this paper. Investigators used the micropuncture technique to sample fluid from nephron sites to measure the ratio of inulin concentrations in this aspirated fluid compared to that in plasma ((TF/P)Inulin) to define the site of water reabsorption in the nephron11). Inulin is used as the marker because it is freely filtered, not reabsorbed, and not secreted in the nephron. Central to our interpretation is the data from fluid aspirated in the early distal convoluted tubule. The (TF/P)Inulin in this nephron was ~6, which indicates that 5/6 of the glomerular filtration rate (GFR) was reabsorbed prior to arriving at this site of micropuncture (i.e., 17% of filtered water was delivered to the early distal convoluted tubule). We did not use inulin in these studies because it must be infused into anesthetized rats, and this would provide non-osmotic stimuli for the release of vasopressin, which would abort the water diuresis. In addition, it is not possible to obtain accurate timed urine flow rates in spontaneously voided urine in rats because bladder emptying is unlikely to be complete. Hence we used the (U/P)Creatinine to assess the urine flow rate and thereby, to indicate how much filtered water was reabsorbed in the nephron. A value of the (U/P)Creatinine that is higher than 6 indicates that water was reabsorbed between this micropuncture site and the final urine.
After close to 300 minutes of maximal water diuresis in 6 rats, the lowest concentration of creatinine in the urine was close to 4.5 mg/dL (400 µmol/L) (Fig. 1), which was 13-fold higher than the concentration of creatinine in plasma [i.e., 0.34±0.01 mg/dL (30±1 µmol/L)] in these rats. This indicates that there is a major site of water reabsorption in the late distal nephron.
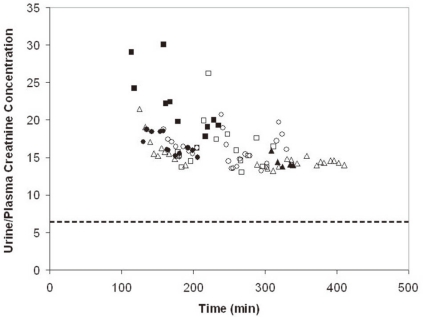 | Fig. 1Effect of a Large Water Diuresis on the (U/P)Creatinine in Rats Given a Large Oral Water Load. The (U/P)Creatinine is depicted on the y-axis and the time of collection of the urine is shown on the x-axis. The results are from 6 separate rats, as indicated by the different symbols. Once the maximum water diuresis began, the (U/P)Creatinine was virtually constant. The (U/P)Creatinine was somewhat more than 2-fold larger than 6 (the dashed horizontal line), which suggests that close to half of the distal delivery of filtrate was reabsorbed downstream for the early distal convoluted tubule, presumably from the inner medullary collecting duct via residual water permeability.
|
In the second protocol, our objective was to measure the osmotic driving force for water reabsorption in the inner medullary collecting duct during a 40 minutes period of water diuresis induced by the ingestion of water that contained sucrose. The urine flow rate in these rats was close to 100 µL/min, which is 300 µL/min/kg body weight (extrapolates to 21 L/day in a 70 kg animal); the concentration of creatinine in the urine was 18 mg/dL (1,600 µmol/L, Table 1). This concentration of creatinine in the urine was ~4-fold higher than in the rats depicted in Fig. 1. Since the duration of the experiment was much shorter than in the first protocol, there may not have been sufficient time for removal all of the AQP2 that were present in luminal membranes of the late distal nephron segments in these rats. In a separate series of 7 rats treated in an identical fashion to those studied during the water diuresis, the osmolality in the excised renal papilla at the first experimental time (10:10 hour) was 774±24 mOsm/kg H2O. In 7 other rats treated in the same fashion, the osmolality in the excised renal papilla obtained at the second time point of this water diuresis (10:45 hour) was 709±19 mOsm/kg H2O (Table 1). These papillary osmolalities were ~600 mOsm/kg H2O greater than the corresponding UOsm (177±36 and 141±35 mOsm/kg H2O, respectively). Hence there was an enormous osmotic driving force for the reabsorption of water in the inner medullary collecting duct at this time (Table 2).


Table 1
Effect of a Large Water Load on the Composition of the Urine and Excised Renal Papilla at Peak Urine Flow Rates
![]()
Table 2
Driving Force for Water Reabsorption Via Residual Water Permeability in the Inner MCD during a Water Diuresis
The major factors that influence the reabsorption of water are the small degree of permeability of the inner medullary collecting duct (MCD) to water and the large osmotic driving force to draw water from its lumen (i.e., the difference in osmolality between the interstitial compartment and the lumen of the inner MCD multiplied by 19.3, the number of mm Hg per mOsm/L). Note that the osmotic driving force is enormous (shown in bold).
![]()
Go to :
Discussion
The principal results in this study were that the (U/P) Creatinine was ~13 during maximal water diuresis (i.e., ~4.5 mg of creatinine/dL (400 µmol/L) in the urine and 0.34 mg of creatinine/dL (30 µmol/L) of plasma, Fig. 1). Of great importance, because the UOsm (141 mOsm/kg H2O, Table 1) was ~ half of the POsm in the second protocol, water did not diffuse to osmotic equilibration in the inner medullary collecting duct. Moreover, the papillary interstitial osmolality was ~600 mOsm/kg H2O was considerably higher than the UOsm (Table 1), which means that the osmotic force to reabsorb water was more than 2-fold greater than implied when the POsm was used in this assessment. The implications of these results will be discussed in terms of residual AQP2 and/or basal or residual permeability of the inner medullary collecting duct to water5).
1. Use of (U/P)Creatinine
Creatinine is produced at a near-constant rate; hence, anesthesia and infusions were not required when this marker of water reabsorption was used. There is little change in its rate of excretion throughout the day12). Moreover, there should be very little change in the secretion or reabsorption of creatinine in individual rats in a short experimental period.
Comparing the (U/P)Creatinine to the (TF/P)Inulin in tubular fluid aspirated from the early distal tubule permits one to calculate how much filtered water was reabsorbed between this micropuncture site and the final urine. Since the (TF/P)Inulin was ~6 in fed rats13, 14), when the (U/P)Creatinine exceeds the 6, filtered water was reabsorbed downstream from this early distal convoluted tubular micropuncture site.
There are two critically important members of the family of water channels in the luminal membranes of the kidney, AQP1 and AQP2. AQP1 are non-regulated and are only present in nephron segments prior to the loop of Henle for the most part15). Of note, AQP1 are also present in the descending thin limbs of the loop of Henle of juxtamedullary nephrons, but not in this nephron segment in the vast majority of nephrons (called the superficial nephrons)16). In fact, only these superficial nephrons are accessible to cortical micropuncture.
Even after a sustained, profound fall in vasopressin levels in plasma, some water is still reabsorbed in the inner medullary collecting duct (called basal or residual water permeability)5). While this might reflect a unique entity, it might also reflect a slower removal of AQP2 from the luminal membrane of the inner medullary collecting duct. The maximum volume of urine during water diuresis is set by the distal delivery of filtrate (equation 1) minus the volume of filtrate reabsorbed via residual water permeability (equation 2).
Distal delivery = GFR - Water reabsorbed in the proximal convoluted tubule (1)
Urine flow rate = Distal delivery - Water reabsorbed downstream to the DCT (2)
There are three features that affect the maximum urine flow rate during water diuresis.
1) Residual or basal water permeability
There basis for suspecting that residual water permeability is small is that water does not diffuse to osmotic equilibrium during water diuresis in normal subjects [i.e., the UOsm is much lower than the osmolality in the medullary interstitial compartment (Table 1)]. It is important to recognize that there is an enormous osmotic driving force for water absorption in this setting (Table 1, 2). On the other hand, we suggested that the volume of water reabsorbed during a water diuresis is very large3, 17) because ~27 L are delivered distally each day, and the maximum urine flow rate during water diuresis is ~20 L/day. Hence ~7 L of water are reabsorbed daily in the inner medullary collecting duct via residual water permeability during an imaginary 24 hour water diuresis (Fig. 2). Although reabsorbing water in the inner medullary collecting duct during water diuresis seems counterintuitive, it may have a useful function (discussed later in this article).
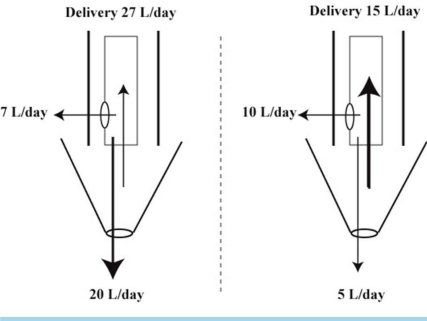 | Fig. 2Contribution of Residual Water Permeability to the Excretion of Water during a Water Diuresis When the Urine Flow Rate is very High and When It Has Decreased. The structures to the left of the dashed vertical line represent a very large water diuresis and the structures to the right of the dashed vertical line represent a modest water diuresis owing to a lower distal delivery of filtrate and a larger reabsorption of water in the inner medullary collecting duct. The upper cylinder of each figure represents the inner medullary collecting duct with bolder outlines to indicate its limited capacity for it to dilate. The inverse triangular structure below it represents the renal pelvis. The arrow below represents fluid that bypasses the retrograde flow and enters the bladder. The numeric values are for illustrative purposes (see text for details).
|
2) Contraction of the renal pelvis
This is an important feature that may affect the maximum urine flow rate during water diuresis. Each time this renal pelvis contracts, some of the volume of fluid currently in this location travels in a retrograde direction up the inner medullary collecting duct18). Nevertheless, not all of this volume enters the inner medullary collecting duct owing to physical constraints (Fig. 2). Therefore, some of this fluid may be reabsorbed via residual water permeability after it enters the inner medullary collecting duct for a second (or third) time. Moreover, this creates a turbulent flow, which may aid both diffusion of water and prolong the contact time. When the volume within the renal pelvis is much smaller, a larger proportion of it may enter the inner medullary collecting duct; hence the volume reabsorbed via residual water permeability can be larger in this setting (right side of Fig. 2).
On the other hand, when the volume of urine that transits through the renal pelvis is very large during a brisk water diuresis, only a small proportion of this delivery may enter the inner medullary collecting duct. Therefore, most of the urine does not have a second opportunity for reabsorption by residual water permeability (left side of Fig. 2). Hence the UOsm should be much lower than the luminal osmolality in the inner medullary collecting duct.
3) Driving force
The magnitude of this force appears to be enormous (i.e., multiply the difference between the papillary osmolality and the UOsm (~600 mOsm/L) by ~19.3 mm Hg, the osmotic driving force when there is a difference of 1 mOsm/kg H2O). Hence this driving force is ~12, 000 mm Hg in the data summarized in Table 2 (19.3 mm Hg×600 mOsm/L).
2. Physiological role of residual water permeability during water diuresis
There is a second imperative following a large intake of water-the luminal fluid in water-impermeable nephron segments and thereby in the final urine should contain as little Na+ as possible. This was especially important in Paleolithic times, as the diet contained very little NaCl19). Moreover, there was an appreciable loss of Na+ and Cl- in sweat20, 21). In addition, females have a loss of Na+ during pregnancy22-24). Most of our important control mechanisms developed in Paleolithic times19, 25); changes induced by our modern diet and activity patterns are unlikely to have sufficient control strength to modify these basic regulatory mechanisms in a meaningful way.
Ensuring that water is reabsorbed in the inner medullary collecting duct during water diuresis has both benefits and potential drawbacks. The major benefit is to diminish the excretion of Na+ and Cl-, which was particularly important in Paleolithic times. The drawback is a diminished ability to excrete water extremely quickly. Nevertheless, this is not likely to be a major problem because the excretion of water is still brisk and the ingestion of sugar-containing fluids in the Paleolithic diet may decrease the rate of stomach emptying, and thereby diminish the rate of absorption of ingested water in the intestinal tract26). There are two major nephron sites where the reabsorption of Na+ and Cl- is stimulated during water diuresis.
1) The medullary thick ascending limb of the loop of Henle
The basic premise is that the reabsorption of Na+ and Cl- in this nephron segment is likely to be regulated because if this region of the kidney were to reabsorb too much Na+, there could be excessive extraction of water from the medullary collecting duct. As a result, the urine flow rate might fall sufficiently to cause high luminal concentrations of sparingly soluble solutes and thereby, increase the risk for kidney stone formation3). In addition, this region of the kidney has a precarious blood supply, which could pose a danger of acute tissue injury when medullary work (active reabsorption of Na+) and thereby, consumption of oxygen is increased27).
Our speculation is that the inhibitory control of the reabsorption of Na+ in the medullary thick ascending limbs of the loop of Henle is mediated by the activity of ionized calcium in the medullary interstitial compartment, which is detected by the calcium sensing receptor on the basolateral membrane of the medullary thick ascending limbs of the loop of Henle3). Hence, a large reabsorption of water from the medullary collecting ducts will lower the activity of ionized calcium in the medullary interstitial compartment (Fig. 3). This, in turn, will augment active reabsorption of Na+ without increasing the risk of precipitation because the initial event was the addition of water to this interstitial compartment. The net result is a small fall in the urine volume, which is the price to pay for having a better composition of the urine and the medullary interstitial compartment during water diuresis.
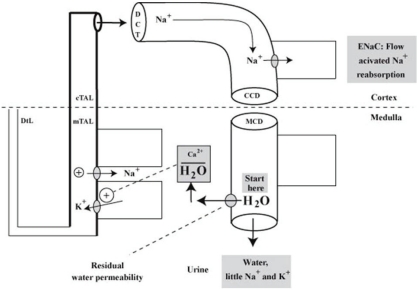 | Fig. 3Desalination of Luminal Fluid in the Medullary Thick Ascending Limb of the Loop of Henle and in the Cortical Distal Nephron. When more Na+ and Cl- are reabsorbed without water, fewer electrolytes will be excreted in a water diuresis. The stimulus begins with a high flow rate in the inner medullary collecting duct (MCD), which leads to more water reabsorption via residual water permeability (shown as a shaded oval near the 'start here' message). As a result, there is a signal (lower concentration of ionized calcium in the outer medullary interstitial compartment), which may increase the reabsorption of Na+ and Cl- in the medullary thick ascending limb of the loop of Henle (mTAL) to begin the desalination process. In the late cortical distal nephron (abbreviated as CCD), flow activation of the epithelial Na+ channels (ENaC) accelerates Na+ reabsorption. cTAL, cortical thick ascending limb; DCT, distal convoluted tubule.
|
2) The late cortical distal nephron
This represents the late distal convoluted tubule, the connecting segment and the cortical collecting duct. Each of these segments has epithelial Na+ channels (ENaC)28). Since flow activation of ENaC has been described29), a high flow rate during water diuresis could stimulate the reabsorption of Na+ in this location (Fig. 3).
3. Clinical example
Our objective in this section is to illustrate the importance of residual water permeability in patients undergoing a water diuresis. A young woman who runs several miles on a regular basis in a warm environment consumes a low salt diet. To avoid 'dehydration', she drinks large volumes of water, but this intake is not driven by thirst. Her main complaint is disturbed sleep because she must void large volumes several times overnight30). The major laboratory findings include a PNa of ~130 mEq/L, urine volume of 5 L/day, and a UOsm ~80 mOsm/kg H2O. Hyponatremia developed because her water intake exceeded her limited ability to excrete water in the urine plus the loss of water and Na+ in sweat31). This low UOsm implies that vasopressin has not acted6).
Using traditional explanations, the diagnosis was polyuria because her daily urine volume that was greater than 2.5 or 3 L. Nevertheless, this is an arbitrary definition of polyuria because it is based on a comparison of this 24-hour urine volume to values observed in individuals who consume a typical Western diet30). This water diuresis is due to primary polydipsia, as her PNa was low.
Using a definition of polyuria that is based on integrative physiology - the 24-hour urine volume should be compared to the urine flow rate in normal adults who have a PNa that is low enough to suppress the release of vasopressin32). For example, following an acute water load that suppresses the release of vasopressin, the urine flow rate is 10 to 15 mL/min4). Extrapolating this flow rate to 1,440 minutes, the expected value would be ~20 L/day. Hence the daily urine volume of 5 L in this patient is very low. Thus she has a form of oliguria because she has a diminished ability to excrete water.
One basis for the oliguria in this patient is a low distal delivery of filtrate secondary to enhanced reabsorption of Na+ and Cl- in her proximal convoluted tubule (equation 1), as she has a low effective arterial blood volume due to ongoing losses of Na+ and Cl- in sweat and a low intake of salt. Evidence to support of this impression is her low osmole excretion rate (80 mOsm/L×5 L/day=400 mOsm/day versus the usual excretion of 600 to 900 mOsm/day in an adult consuming a typical western diet).
For simplicity, we shall assign a value of 15 L/day to her distal delivery. There would need to be a powerful stimulus to reabsorb an extra 12 L of filtrate and thereby 1,800 mEq more Na+ and Cl- in the proximal convoluted tubule. It is likely that she did have a lower effective arterial blood volume, but its degree was not large enough to decrease her distal delivery of filtrate by much larger than to 15 L/day, as there were no signs of this on physical examination and she was able to perform vigorous exercise on a regular basis. If this interpretation were valid, she would need a second mechanism to have such a low urine flow rate in the absence of actions of vasopressin. A mechanism that could contribute to the low urine volume is the reabsorption of more water in the inner medullary collecting duct via residual water permeability18). In this context, as the urine flow rate declines during a water diuresis, a larger proportion of the potential urine undergoes retrograde flux from the renal pelvis and this presents a greater opportunity to reabsorb more of the distal delivery in the inner medulla for the reasons mentioned above (Fig. 2).
4. Concluding remarks
Two facts are well known in a water diuresis. First, vasopressin must be absent, which implies that AQP2 are not present in the luminal membranes of the late distal nephron. Second, the upper limit on the rate of excretion of water is set by the distal delivery of filtrate. Our objective in this study was to provide information about other factors that might limit the excretion of water during a water diuresis. The finding of a lower urine flow rate than the estimated distal delivery of filtrate adds support for the presence of this phenomenon. We speculated that the reabsorption of this water could stimulate the reabsorption of Na+ and Cl- in the medullary thick ascending limb of the loop of Henle3), which contributes to the desalination of the final urine. The potential clinical importance of this physiology was illustrated in a case example.
Go to :
 XML Download
XML Download