

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The kidneys are responsible for the regulation of body water and electrolyte metabolism. Thus, understanding of the underlying mechanisms for renal water transport is critical. Water permeability along the nephron has already been well characterized in the mammalian kidney1). Approximately, 180 L/day of glomerular filtrate is generated in an adult human, more than 80-90% of the glomerular filtrate is constitutively reabsorbed by the highly water permeable proximal tubules and descending thin limbs of Henle's loop. The ascending thin limbs and thick limbs are relatively impermeable to water, but the collecting ducts are important, because they are the chief site of vasopressin-regulated water reabsorption2). Aquaporins, water channel proteins, are chiefly involved in the collecting duct water transport3). In particular, aquaporin-2 (AQP2), the vasopressin-regulated water channel protein in the collecting duct principal cells, plays a critical role in water reabsorption by both short-term regulated trafficking of AQP2-expressing vesicles to the apical plasma membrane4) and long-term regulation of AQP2 protein abundance5). This process produces concentrated urine and is essential for regulation of body water metabolism6). In contrast to the well-established signaling pathways for the vasopressin-regulated AQP2 trafficking and up-regulation of AQP2 expression, the underlying mechanisms for AQP2 endocytosis and intracellular degradation of AQP2 protein are unclear. So far, two hormones (prostaglandin E2 and dopamine) cause AQP2 internalization independent of S256 dephosphorylation7,8). Similarly, in Madin-Darby Canine Kidney (MDCK) cells, activation of protein kinase C by the phorbol ester 12-tetradecanoylphorbol-13-acetate (TPA) results in the internalization of AQP2 without affecting AQP2 phosphorylation at S256 or inducing any other phosphorylation of AQP29).
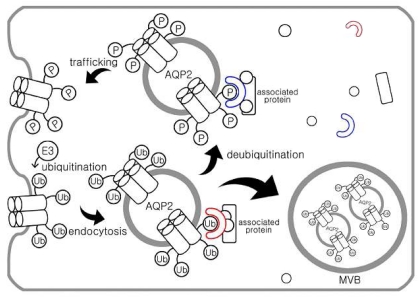
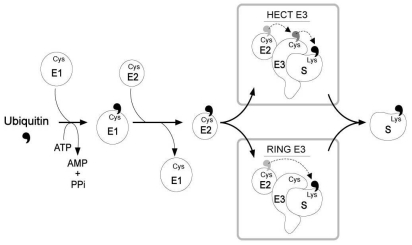
Two major pathways for intracellular protein degradation are the ubiquitin-proteasome pathway and lysosomal proteolysis. Consistent with this, a recent study demonstrated that ubiquitination is important for endocytosis and lysosomal degradation of AQP2 (Fig. 1). The results revealed that AQP2 is polyubiquitinated at the plasma membrane on a single residue (K270), resulting in the internalization of AQP2, transport to multivesicular bodies and subsequent proteasomal degradation10). Ubiquitin (Ub) is a 76-amino acid protein, which is covalently attached to the lysine residue of the substrate proteins. AQP2 has three putative attachment sites (cytosolic lysine residues) for ubiquitin at positions 228, 238, and 270. They made AQP2 mutants having one, two, or all three lysines replaced by arginines and these mutants were expressed in MDCK cells10). Ubiquitination analyses demonstrated that all mutants having the K270R replacement were not ubiquitinated, whereas all other mutants revealed the similar intensity of ubiquitination as wild type AQP210). Therefore, these data indicate that K270 is the only substrate for ubiquitination in AQP2. Activation and attachment of Ub to a target protein is mediated by the action of three enzymes (i.e., E1, E2, and E3) which involves the formation of thioester intermediates with the C-terminal glycine residue of Ub11). The E1 (Ub-activating enzyme) activates the C-terminus of Ub in an ATP-dependent manner, and both E2 (Ub-conjugating enzyme) and E3 (Ub-protein ligase) are involved in the attachment of Ub to a target protein specifically through the ε-amino group of a lysine residue12-14)(Fig 2).
In particular, E3 Ub-protein ligases are known to have substrate specificity, and hence they play an important role in determining the selectivity of Ub-mediated protein degradation15,16). For example, epithelial sodium channel (ENaC), expressed in the collecting duct principal cells in the kidney, is composed of α-, β-, and γ-subunits that contain PY motifs recognized by WW domains of Nedd4, a member of the homologous to E6-associated protein C-terminus (HECT) E3 Ub-protein ligase family17,18). The magnitude of ENaC-mediated sodium transport is determined by the open probability of the channels and the number of channels in the apical plasma membrane of collecting duct principal cells19). The number of channels in the apical plasma membrane is regulated by the rate of ENaC insertion and the rate of removal from the membrane. The carboxyl-terminal domain of all three subunits is intracellular and contains a proline-rich motif19). Mutations or deletion of this proline-rich motif in the β- and γ-subunits (i.e., Liddle's syndrome) inhibits the binding of a specific E3 ubiquitin ligase, Nedd4-2, to the channel, thereby impeding ubiquitin conjugation of the channel subunits, and maintaining the ENaC expression in the membrane19). Thus, ubiquitin conjugation would regulate the rate of ENaC turnover in the kidney collecting duct principal cells and the rate of ENaC degradation is apparently controlled by Nedd4-2 activity. However, no studies have identified the specific E3 Ub-protein ligases for AQP2 endocytosis and intracellular degradation.
For identifying proteins interacting with AQP2 C-terminus, several methodological approaches could be exploited, e.g., a yeast-two-hybrid assay, in vitro phage display technique, transcriptomome analysis, and liquid chromatography-tandem mass spectrometry (LC-MS/MS) proteomic analysis20-24). Recently, van Balkom et al.21) isolated lysosomal trafficking regulator-interacting protein 5 (LIP5) by a yeast-two-hybrid assay and found that LIP5 interacts with the proximal carboxy-terminal tail (L230-D243) of AQP2 in vitro, but not with AQP3 or AQP4. Interestingly, LIP5 binding occurred independent of the state of Ser256 phosphorylation or Lys270 ubiquitination. Knockdown of LIP5 in mouse renal cells (mpkCCD) reduced the phorbol ester-induced degradation of AQP2 approximately two-fold, suggesting that LIP5 is likely to play a role in protein sorting to multivesicular bodies21).
On the other hand, we recently identified several E3 Ub-protein ligases putatively associated with AQP2 regulation in the kidney of a rat model with 1-deamio-8-D-arginine vasopressin (dDAVP) withdrawal for periods after 5 day-dDAVP infusion (not published). In order to do this, 1) rats were infused with vehicle or dDAVP for 5 days, or dDAVP stimulation was withdrawn for periods after 5 day-dDAVP infusion for inducing AQP2 endocytosis and degradation; 2) whole kidney protein samples, and plasma membrane (PM)- or intracellular vesicle (ICV)-enriched membrane fractions were analyzed by AQP2 immunoblotting and kidney sections were labeled with anti-AQP2 antibody for examining the changes of intracellular AQP2 expression during dDAVP withdrawal; 3) PM- or ICV-enriched membrane fractions of whole kidney immunoisolated with anti-Ub antibody were subjected to AQP2 immunoblotting; 4) immunoisolated PM- or ICV-enriched membrane fractions with anti-AQP2 antibody or anti-Ub antibody were trypsin-digested and subjected to LC-MS/MS analyses; 5) total RNA isolated from inner medulla was subjected to gene chip analysis using Affymetrix GeneChip® Rat Gene 1.0 ST array; 6) mRNA expression of identified E3 ligases were examined by RT-PCR in whole kidney and inner medulla; and 7) time course of changes in the expression of selected E3 ligases was examined by immunoblotting in rats with vehicle treatment, dDAVP-treatment for 5 days or 3 h-dDAVP withdrawal after 5 day-dDAVP infusion. Both transcriptome analysis and liquid chromatography-tandem mass spectrometry (LC-MS/MS) proteomic analysis revealed five common isoforms of E3 ligases (UBR4, UHRF1, NEDD 4, BRE1B, and Cullin-5), which are putatively associated with dDAVP-induced AQP2 regulation. Further studies including knockdown study with siRNA transfection are needed to find out whether these identified E3 ligases play a role in Ub-conjugation to AQP2 and thereby AQP2 endocytosis and intracellular degradation by the ubiquitin-proteasome pathway.
 XML Download
XML Download