

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
As a result of both improved technology and understanding of neurological diseases, there has been a significant increase in the application of restorative functional neurosurgical techniques to treat neuropsychiatric disorders. Among these newer surgical therapies, electrical stimulation of specific subcortical brain nuclei, known commonly as deep brain stimulation (DBS), has become an increasingly popular alternative to pharmacological treatment alone. At this time, more than 75,000 people have been successfully implanted with DBS devices worldwide, and this number is expected to expand rapidly. DBS is now approved by the US Food and Drug Administration (FDA) and is in routine clinical use for treatment of, Parkinson's disease (PD),1 essential tremor (ET),2-4 dystonia5 and obsessive compulsive disorder (OCD).6,7 Furthermore, there is growing evidence for the use of DBS in the treatment of disorders such as depression,8-10 epilepsy,11-13 Tourette syndrome (TS),14 and chronic pain.15,16 Here, we review current and emerging evidence for the role of DBS in the management of a range of neurological and psychiatric conditions, discuss the potential mechanism of action, and explore the future application of this exciting technology.
Parkinson's Disease
PD is associated with a severe loss in function of dopaminergic neuronal cells within the substantia nigra pars compacta which project to the striatum, a major component of the basal ganglia. Progressive degeneration of these nigrostriatal projections leads to motor symptoms such as tremor, rigidity, bradykinesia, and postural instability. While PD patients have benefited significantly from the development of new pharmacological treatments used in combination with the traditional drug levodopa, many of these therapies have been either only partially effective or poorly-tolerated over the long course of the disease.17,18 In addition, these pharmacological therapies are also associated with serious and debilitating motor complications, such as dyskinesias.19 However, significant advances in stereotactic and functional neurosurgical techniques over the last 15 years in response to these shortcomings have led to new strategies in the treatment of advanced PD using electrical stimulation.20-22
Although the therapeutic efficacy of DBS in PD has been well-established,23,24 results from the multicenter, open-label PD SURG trial in the UK have shown that DBS plus medical therapy improves patient self-reported quality of life significantly more than best medical therapy alone.25 Importantly, these findings were accompanied by clinically meaningful differences on the Unified Parkinson's Disease Rating Scale (UPDRS), including substantially improved patient ratings of the frequency and severity of dyskinesias and "off" periods.
Today, the most common target for DBS in PD is the subthalamic nucleus (STN) as this ameliorates the cardinal symptoms of bradykinesia, rigidity and tremor.24,26,27 In cases of PD where the patient's main complaints do not involve bradykinesia or rigidity, other anatomical targets for DBS may be appropriate. For example, DBS of the ventralis intermedius (Vim) thalamus, which is common for treating ET, is also an effective target for patients with tremor-dominant PD.28-32 As mentioned, studies have also demonstrated significant overall improvements in PD patients treated with DBS of the globus pallidus interna (GPi).33-42 Although improvements in both gait and posture had been shown,37,43 the main utility of GPi DBS was seen to be in the reduction of dyskinesias often seen with long-term levodopa treatment.36-42,44 However, the results of a recent multicenter, randomized, blinded trial showed that both GPi and STN DBS result in similar improvements in part III (motor subscale) of the UPDRS at 24 months.45 Patients who underwent STN DBS subsequently required lower doses of dopaminergic agents, and the level of depression (as measured by the Beck Depression Inventory II) worsened slightly after STN stimulation but improved slightly after GPi stimulation, although the reported differences are unlikely to be clinically important. Recently, interest has also grown in low frequency stimulation of the pedunculopontine nucleus for gait disorders/freezing and postural instability in PD, with two small double-blinded studies showing a reduction in falls at 1-year.46,47
Essential Tremor
ET is characterized by rhythmic, involuntary movements commonly affecting the arms, head and voice.48 It is by far the commonest movement disorder, with a prevalence of up to 5.5% in those 65 and over.49 Early promising results for DBS in the treatment of ET50 were replicated and eventually led to a direct comparison study of thalamotomy and DBS of the Vim thalamus for treatment of tremors, including ET.51 In this study, Schuurman et al.51 reported a significant improvement of tremor in both treatment groups, but a greater reduction in the DBS group, who also experienced fewer side effects and a better functional outcome. The marked improvement seen with DBS of the Vim thalamus for upper limb tremor also appears to be maintained during long-term follow-up.52
Dystonia
Dystonia refers to a broad group of conditions with differing aetiologies, which involve abnormal muscle spasm and posturing. In general, they may be primary, associated with a genetic predisposition such as the torsion dystonia 1 (DYT1) gene, or secondary to other causes such as metabolic disorders, drug intake (e.g., tardive dyskinesia), or brain injury (e.g., stroke).55 Dystonias are also classified as generalized, segmental (affecting two adjacent body parts) or focal (e.g., cervical dystonia/spasmodic torticollis, blepharospasm, and writer's cramp) according to the pattern of involvement.
Most evidence for DBS in dystonia highlights the effectiveness of targeting the GPi in primary generalised dystonia56,57 and cervical dystonia. There is also increasing evidence for bilateral GPi DBS in patients with tardive dyskinesia,58-60 a condition which is notoriously difficult to treat medically. Secondary dystonias tend to benefit less from DBS of the GPi, and this may be a consequence of the wide range of aetiologies and brain areas potentially involved.
DBS for Psychiatric Disorders
As highlighted by Kuhn et al.61 the idea of using DBS to treat psychiatric disorders stems from several observations and developments: 1) some PD patients treated with DBS developed psychiatric adverse effects or experienced improvement of a co-morbid psychiatric disorder; 2) lesion procedures for intractable psychiatric disorders have yielded positive results, but were avoided because of their severe adverse effects; 3) identification of potential anatomical targets for DBS in psychiatric disorders has improved due to a growing body of functional neuroimaging studies. The role of neuromodulation in the treatment of refractory psychiatric disorders has been extensively reviewed, including by our own group62 and more recently by others.63 Here, we will only provide an overview of current thinking about the role of DBS in treatment-resistant cases of depression, OCD and TS.
Depression
In the United States, major depression may affect almost 1 in 5 people64 and up to 20% of patients fail to respond to first-line pharmacological interventions.65 Early studies have implicated the subgenual cingulate cortex (Cg25) in acute sadness and antidepressant effects,66,67 and a decrease in Cg25 activity has been associated with immediate clinical response to a number of antidepressant treatments including serotonin reuptake inhibitors,68 electroconvulsive therapy,69 transcranial magnetic stimulation,70 and ablative surgery.71 As such, early research by Mayberg et al.9 focused on bilateral Cg25 white matter (Cg25WM) DBS for treatment-resistant depression and resulted in the publication of a preliminary report of six patients9 and a final report of twenty patients.72 In the final report, after 6 months 12/20 patients had a reduction of at least 50% in the 17-item Hamilton Rating Scale for Depression (HRSD-17) score and 7 patients met criteria for remission (HRSD-17 ≤7). Positron emission tomography (PET) of cerebral blood flow (CBF) in the pilot study (5/6 patients) showed increased Cg25 CBF and decreased CBF in lateral prefrontal and anterior cingulate cortices relative to controls, which reversed with stimulation. Of the additional 14 patients in the final study, 18-Fluoro-deoxyglucose-PET (FDG-PET) results from 8 patients responsive to DBS similarly showed widespread changes in cortical and limbic metabolic activity, including increased activity in lateral prefrontal cortex and Cg25WM, but a reduction in Cg25 grey matter.
Other potential targets relate to areas which the Cg25 sends and receives its projections: the nucleus accumbens (NAcc) [also referred to as the ventral striatum (VS)] and limbic cortical loop.73 The NAcc forms an interface between emotional, limbic and motor neuronal circuits and plays a vital processing role in the experience of reward and hedonistic stimuli.61 In a study by Schlaepfer et al.10 3 patients with treatment resistant depression underwent bilateral NAcc DBS. Within one week of stimulation onset the HRSD-24 decreased by an average of 42% and demonstrated that stimulation was inversely correlated with depression. Interestingly, while FDG-PET after 1 week of NAcc DBS only showed activation of dorsal prefrontal and cingulate cortices (with no change in Cg25 activity), subsequently published expanded data for the first 10 patients74 showed decreased metabolism in both the Cg25 and prefrontal regions on FDG-PET taken at 6 months. In this study, 5 patients reached 50% reduction in HRSD-28 at 1 year and anxiety was reduced in the whole group, but to a greater degree in the responders.
Previously, studies targeting the ventral anterior internal capsule (VC)/VS treatment-resistant OCD patients have also shown improvements in depressive symptoms.7,75-80 As such, Malone et al.81 attempted bilateral VC/VS DBS in 15 patients with treatment-resistant depression. They found that the proportion of patients with at least 50% reduction in HRSD-24 was 47% at 3 months, 40% at 6 months and 53% at last follow up, while remission rates with HRSD-24 were 20% at 6 months and 40% at last follow up.81
Obsessive Compulsive Disorder
OCD is characterized by recurring, anxiety-provoking thoughts (obsessions) and repetitive behaviors (compulsions), and affects approximately 2% to 3% of the general population.62,82 The FDA granted DBS a humanitarian device exemption for medically refractory OCD in 2009 on the basis of several clinical trials showing positive findings. As such, the main targets currently used for DBS in OCD are the VC/VS,79 the shell region of the NAcc, and the STN.
Early studies showed that bilateral VC/VS DBS had a beneficial impact on OCD symptoms, but required unusually high stimulation amplitudes (5.0-10.5 V), suggesting that perhaps the therapeutic target was adjacent to, but not at the electrode site. Taken together, the high stimulation amplitudes and electrode site (the ventral edge of the internal capsule, where it abuts the NAcc) has prompted suggestions that functional blocking of NAcc activity may be underlying the symptomatic improvement.83 Indeed, a study of unilateral right NAcc DBS in 4 patients with refractory OCD showed near complete recovery in 3 out of 4 patients using lower stimulation amplitudes (2.0-6.5V).83 Recently, in a double-blind sham-controlled crossover study of unilateral right NAcc DBS in 10 patients with treatment-resistant OCD, Huff et al.84 reported a full response [>35% reduction in Yale-Brown Obsessive Compulsive Scale (Y-BOCS)85] in one patient and partial responses (25-35% reduction in Y-BOCS85) in 4 other patients at 1 year. Despite a different target and only using unilateral stimulation, these results are broadly consistent with earlier studies7,78,86 where about 50-60% of patients acquired a ≥25% reduction in Y-BOCS score within one year of DBS. While the majority of patients in these earlier studies had full responses, Huff et al.84 suggest that this may be due to differences in stimulation protocol, anatomical target, use of bilateral stimulation, and effectiveness of blinding.
In patients with OCD who underwent STN DBS for their PD symptoms, studies have also reported unintentional psychiatric benefits.87,88 In view of this, Mallet et al.89 performed a randomized double-blind crossover study of bilateral anteromedial STN DBS for treatment-resistant OCD, with two parallel groups of 8 patients undergoing two sequential 3-month blinded phases ("stimulation on" followed by "stimulation off" vs. "stimulation off" followed by "stimulation on"). This study found a significant reduction in the Y-BOCS score after the on-stimulation phase compared to after the sham-stimulation, but urged caution after observing 15 serious adverse effects in 11 patients (including one intracerebral haemorrhage and 2 infections).
Tourette Syndrome
TS is characterized by repetitive, stereotyped, involuntary movements and vocalizations called tics.62 These can be simple motor tics (e.g., eye blinking, facial grimacing, shoulder shrugging, repetitive throat-clearing), complex tics (combinations of movements) and vocal tics such as coprolalia (uttering swear words) or echolalia (repeating the words or phrases of others). The early symptoms of TS are generally noticed in childhood, with an average age of onset of 7 to 10 years.90 Although in most cases the disorder is self-limiting or amenable to treatment, some patients have an intractable form which may be helped by DBS.91
In 1999, DBS was trialed as a new approach for intractable TS.92 Maciunas et al.14 conducted the first prospective double-blind crossover trial of DBS in five adults with TS, utilizing bilateral stimulation of the centromedian-parafascicular thalamic nuclei (CM-Pf). In the initial 4-week blinded phase patients spent each week in one of 4 electrode states [no stimulation; unilateral (right-sided) stimulation only; unilateral (left-sided) stimulation only; bilateral stimulation], followed by 3 months in the bilateral stimulation state. During the initial blinded phase, a >50% reduction in tics was seen in 3/5 patients during the bilateral stimulation week (versus no stimulation) as measured by modified Rush Video-Based Rating Scale and Yale Global Tic Severity Scale (YGTSS), which fell to 2/5 patients after 3 months of open stimulation. In another double-blind, randomized crossover trial 3 patients with medically refractory TS were implanted with DBS electrodes in the CM-Pf and GPi bilaterally (i.e., 4 electrodes per patient).93 In the 8-month blinded phase, patients experienced 2 months each of CM-Pf, GPi, combined CM-Pf/GPi, and sham stimulation. Interestingly, the reduction in the YGTSS was greater during bilateral GPi stimulation alone (about 78%) compared to bilateral CM-Pf stimulation (about 45%), and even compared to simultaneous CM-Pf/GPi stimulation (about 60%).
In another uncontrolled study, 18 cases of TS underwent DBS placed bilaterally in the CM-Pf and ventralis oralis complex of the thalamus.94 Although follow up ranged from 3-18 months, the average YGTSS fell by about 70% after 12 months. The same group has recently published the 2-year outcomes for 15 of the original 18 patients, which showed a sustained reduction in tic severity (about 53%), as well as improvement in obsessive-compulsive symptoms, anxiety symptoms, depressive symptoms, and subjective quality of life.95
Epilepsy
Approximately 35% of epileptic patients experience refractory disease unresponsive to antiepileptic drugs.96 While some may benefit from resective surgery, patients who have seizures arising from eloquent cortex, or which are multifocal, bilateral, or generalized, are not candidates for resective surgery and can be considered for alternative therapy.97 In this context, DBS aims to reduce seizures by modulating subcortical systems which can remotely control seizure generators. Indeed, previous studies have looked at DBS applied to a number of subcortical targets, including the cerebellum, various thalamic nuclei, and several structures of the basal ganglia system (reviewed by Kahane and Depaulis97). However, the recent publication of the results from the multicenter, double-blind, randomized SANTE trial of bilateral DBS of the anterior thalamic nuclei in patients with treatment-resistant partial and secondarily generalized epilepsy represents a significant step. This study showed a 56% reduction in mean seizure frequency at 2 years (n=102), with 54% of patients having at least 50% reduction in seizure frequency.98 While complication rates were modest, 2 participants had transient stimulation-related seizures and during the initial 3 month blinded phase participants in the sti-mulated group were more likely to report depression and memory problems as adverse effects.
Chronic Pain
DBS for chronic pain has been used since the 1950s, when the caudate and thalamus were targeted in patients with chronic pain. However, the lack of conclusive results from multicentre trials99 undertaken thus far has meant that DBS for pain is performed on an off label basis. In a meta-analysis of 7 studies published from 1977-1997, which included 424 patients undergoing DBS to the PAG/PVG, internal capsule, and thalamus, Bittar et al.16 found a long term success rate of 79% for PAG/PVG compared to only 58% for sensory thalamic stimulation, but 87% when PAG/PVG was combined with sensory thalamic/internal capsule stimulation. Interestingly, this is in contrast to findings of a recent study of 21 patients receiving either ventrocaudalis thalamic nucleus (Vc) or combined Vc and PAG/PVG DBS, where of the 5 patients who experienced long-term pain relief after DBS, 4 had received Vc only DBS.100 The analysis by Bittar et al. also found that patients with failed back surgery syndrome (FBSS) had better outcomes than those with post-stroke pain, phantom limb pain or peripheral neuropathic pain, and that nociceptive pain responded to DBS better than neuropathic pain. More recently, in a study of 56 patients with various neuropathic and mixed nociceptive/neuropathic pain syndromes undergoing DBS of the PVG/PAG and either VPL or VPM, the best results were again seen in patients with FBSS, while poorer outcomes were seen for dysesthesia dolorosa, phantom limb pain and central pain syndromes (spinal cord injury and post-stroke pain).15 Interestingly, smaller studies by a group in Oxford, UK have had significantly better outcomes for phantom limb pain,101 neuropathic pain102 and central pain. Indeed, in a trial of 15 patients with post-stroke pain who underwent DBS of the PVG and VPL, it was initially successful and devices internalized in 12 patients exhibited a nearly 50% reduction in visual analogue pain score over a follow-up period of 27 months.103
The role of DBS in treating neuropathic and other facial pain syndromes has also been studied. Franzini et al.104 targeted the ipsilateral posterior hypothalamus for DBS and reported long-term pain relief in 5 patients with treatment-resistant cluster headache (CH). Subsequently, Schoenen et al.105 studied ipsilateral ventroposterior hypothalamic DBS in 6 patients with intractable CH. Surgery was successfully completed in 4 patients and, of these, 2 were pain-free and 1 had <3 attacks per month, but another only had transient remissions. Long-term results of continuous posterior inferior hypothalamic stimulation in 16 chronic drug-refractory patients with CH showed that 13 patients were persistently pain-free or almost pain-free, and the other 3 are improved at a mean follow-up of 23 months.106 More recently, Green et al.107 studied DBS of the PAG/PVG and/or VPM in 7 patients with a range of neuropathic cephalgias of various aetiologies and demonstrated >50% improvement in their pain scores.
DBS Surgical Procedures
All approaches to DBS surgery broadly combine stereotactic technique with detailed image guidance. Commonly, a stereotactic head frame is placed on the patient under local anesthesia and magnetic resonance imaging (MRI) is performed to identify the anterior commissure, posterior commissure, and the mid-commissural point. Well-established Cartesian (x, y, and z) target coordinates, relative to the mid-commissural point, are used for planning electrode placement. Stereotactic target coordinates are discerned from computer software that merges the MRI of the patient's brain with a brain atlas. At our institution, we routinely utilize gadolinium contrast in our pre-operative MRI with a head frame to visualize the location of blood vessels which, in addition to the ventricles, can then be avoided in the planned electrode trajectory.
Once imaging has been completed and a safe trajectory established, the patient is returned to the operating room where, under sterile conditions and local anesthesia, surgery commences. One or more burr holes are placed in the skull at the predetermined entry points. During surgery microelectrode unit recordings are used to verify a trajectory using region-specific neural activity as functional landmarks.108,109 If suitable results are not obtained, another tract is chosen, and the recording procedure is repeated. Once a trajectory is verified, the microelectrode is withdrawn and the stimulating electrode implanted and test stimulation conducted in the awake patient using a temporary external stimulator. This enables the patient to give verbal feedback confirming the absence of any unwanted side effects from stimulation (e.g., paraesthesias suggesting current spread to the somatosensory thalamus) and provides the surgeon with an opportunity to relocate the electrode if side effects do occur. Confirmation of final electrode position is usually performed first with intraoperative fluoroscopy then postoperative MRI or computed tomography. Once complete and trial stimulation is deemed successful, the implanted stimulating electrode is secured to the skull and connected to a pulse generator that is subsequently implanted subcutaneously, inferior to the clavicle.
As such, DBS surgical procedures can be quite lengthy, requiring the awake patient to undergo many difficult hours of electrophysiological recordings that is necessary to confirm the anatomy scanned by MRI. With each pass of the recording microelectrode there is an increased risk of intracranial hemorrhage.110 Even with this extensive and sometimes precarious implantation procedure, stimulating electrodes may be misplaced, an event associated with side effects such as depression111,112 and even suicidal ideation.112 The state of DBS technology, the surgical procedures, and post-surgical imaging have remained largely static since their inception and remain arduous and time consuming for both patient and practitioner. As such, obtaining optimal lead placement with minimal lead penetrations is of paramount importance and the availability of more refined anatomical information to enable more precise placement of DBS electrodes, together with utilization of real-time electrophysiological and neurotransmitter measurements during surgery, may address many of these critical issues. However, it is clear that the framework for the development of the next generation of DBS devices and surgical approaches is dependent on our understanding of its mechanism of action.
DBS: Potential Mechanisms of Action
Despite the bourgeoning application of DBS in clinical practice, an understanding of the mechanism(s) underlying its clinical effect is incomplete. The similar effectiveness of STN DBS and ablative surgery targeting the STN (subthalamotomy) initially led to the idea that DBS acted to silence pathologically hyperactive neurons113-116 and this was supported by electrophysiological studies.117,118 Paradoxically, more recent studies have reported the activation of STN output nuclei during DBS.119 However, mathematical modeling has reconciled these two finding by suggesting that, because of dissimilar excitability of neural elements, soma inhibition and axonal activation are both expected at the DBS electrode site.120,121 This axonal activation hypothesis, which has now come to dominate current thinking122-125 proposes that DBS evokes changes in neural activity and neurochemical transmission in interconnected structures within the basal ganglia complex that ultimately underlie clinical benefit. Unfortunately, our present understanding of these distal effects of STN DBS remains far from complete, in large part because of the technical difficulties in combining available modalities for the global assessment of neural activity with those for the detection of specific neurochemicals.
Consistent with the axonal activation hypothesis, electrophysiological recordings during STN DBS have shown increased activity in STN target neurons in the GPi and globus pallidus externa (GPe)126-128 and the substantia nigra pars reticulata and pars compacta (SNr and SNc, respectively).129-131 While definitive, the downside of the electrophysiological approach is that targets must be selected a priori and few targets can be evaluated concurrently. In contrast, brain imaging techniques are ideal candidates for simultaneous global assessment of neural activity during STN DBS.
Several clinical studies utilizing PET, H215O-PET, and FDG-PET support the axonal activation hypothesis of STN DBS.132 PET and H215O-PET record changes in regional cerebral blood flow (rCBF),133,134 while FDG-PET measures regional cerebral glucose metabolism.135 Both rCBF and metabolic activity are considered to reflect altered local neuronal activity or altered input into the region of measurement.136 In a total of 70 PD patients using PET133,137 and H215O-PET132 in the resting state, studies have found that with clinically effective STN DBS there are similar increases in activity in the globus pallidus, thalamus, and SN and decreases in activity in the pre-motor and supplementary motor areas, including the primary motor cortex. Zhao et al.138 using FDG-PET in seven PD patients also found similar increases in activity in basal ganglia structures, including caudate nucleus and putamen (striatum). Taken together, these PET results suggest that the net effect of STN DBS is to increase the activity of STN output, supporting the axonal activation hypothesis.
Clinical studies utilizing functional MRI (fMRI) have also supported the axonal activation hypothesis of STN DBS. The fMRI brain imaging protocol measures blood-oxygenation-level dependent (BOLD) contrast139 that provides in vivo real-time anatomic maps of blood oxygenation in the brain under normal physiological conditions.140,141 In the first attempt to utilize 1.5 Tesla (1.5 T) fMRI in four PD patients during STN DBS, Jech et al.142 showed BOLD signal activation in structures in the basal ganglia complex such as the globus pallidus, thalamus, SN, and cortical structures that included premotor cortex and dorsolateral prefrontal cortex. In a more recent fMRI study examining the effects of STN DBS, Philips et al.143 implanted five PD patients with bilateral DBS electrodes. These investigators reported that BOLD signal activation was seen in the ipsilateral basal ganglia, typically in the caudate nucleus, putamen and GP in all subjects and ipsilateral thalamus in six of the electrodes tested. In another fMRI study of one subject the electrode on the left was within the ventral STN, whereas the right electrode was in the dorsal STN.144 The left STN DBS primarily showed increases in premotor and motor cortex, thalamus, putamen, and cerebellum, as well as decreases in sensorimotor/supplementary motor cortex while the right DBS showed similar but less extensive change in motor regions and unique increases in prefrontal cortex, anterior cingulate, thalamus, caudate nucleus, and brainstem. Where STN DBS effects were specifically examined in PET and fMRI studies, they clearly demonstrate that increases in basal ganglia network activity (including in the striatum) are consistent with the axonal activation hypothesis, but the question of which neurotransmitter systems are responsible remains unresolved.
The dopamine hypothesis
The cardinal symptoms of PD (akinesia, rigidity and tremor) are associated with severe nigrostriatal dopaminergic denervation145 and levodopa, a biochemical precursor to dopamine and the mainstay of PD treatment, is thought to act by increasing endogenous dopamine synthesis and release.146-148 Bilateral STN DBS reverses the cardinal motor symptoms in PD patients,26,44 and decreases or eliminates the need for levodopa.149,150 However, the hypothesis that DBS of the STN contributes to symptom relief in PD by activation of surviving nigrostriatal dopaminergic neurons, resulting in dopamine release and resumption of target cell control in the striatum, is still controversial. Indeed, it is entirely possible that STN DBS could be altering neuronal circuits downstream of striatal dopamine release to provide therapeutic benefit.
Most basic (animal) studies using in vivo microdialysis, which physically removes analyte from brain extracellular fluid for ex vivo analysis, do not report an increase in striatal dopamine release during high frequency stimulation (HFS) of the STN in intact rats or the 6-hydroxydopamine (6-OHDA)-lesioned rat model of PD.151-153 Additionally, Windels et al.154,155 have shown in rats that STN DBS significantly increased glutamate and GABA release in the GPe and SNr, respectively. However, the relatively large size of microdialysis probes have been shown to disrupt tissue in the immediate vicinity of the probe resulting in underestimations of extracellular dopamine levels compared to alternative measurement techniques that utilize chemical microsensors.156-158 As such, approaches other than microdialysis will be necessary to assess striatal dopamine release during STN DBS. Indeed, chemical microsensors, which offer a smaller probe (5-10 µm versus 200-400 µm diameter for microdialysis probes), have shown dopamine release in the striatum evoked by STN DBS in the intact and 6-OHDA rat model159-161 and in an intact large animal (pig) model.162 These latter findings are important on several levels. For example, striatal dopamine release during STN DBS has been difficult to establish with microdialysis,151-153,163 with one exception,164 a result that underscores the need for application of much smaller microsensors in neurochemical assessments of the effects of STN DBS on distal neurotransmission.
Several in vivo PET studies have also failed to demonstrate significant displacement of the dopamine receptor ligand [11C] raclopride despite significant improvements in motor performance following STN DBS,165-168 potentially suggesting that terminal dopamine release does not underlie its anti-Parkinsonian effects. However, PET scanning with raclopride has relatively poor temporal resolution and requires an increase of greater than 90% of baseline measures in order to detect a change in dopamine efflux.166,169 Additionally, it has been suggested that adaptive changes in dopamine receptor populations (e.g., D2 receptor internalization and/or recycling) occurring over long-term STN stimulation may interfere with PET quantification of dopamine release in these patients.170 However, the fact that DBS of the STN is most effective in PD patients who respond well to levodopa171 and contraindicated for those who do not,172 suggests that effective DBS requires endogenous dopamine production. Additionally, the observations that DBS elicits dyskinesias that resemble those seen with levodopa excess26 and that, like levodopa, it contributes to impulsivity (a behavior thought to be dopamine-mediated)173 are also consistent with activation of surviving dopaminergic neurons by DBS. Thus, whether STN DBS improves PD symptoms via the release of dopamine remains an important but unanswered question.
Adenosine, glutamate, and glia
Another putative neurochemical that may be of importance to STN DBS mechanisms is adenosine. Proposed as a chemical mediator of thalamic DBS for the treatment of ET,174 adenosine release can be measured in the striatum with chemical microsensors during electrical stimulation in the vicinity of the nigrostriatal dopaminergic tract.175,176 Importantly, increases in extracellular adenosine appear to match elevations in CBF resulting from increases in neural activity.177 Indeed, we have demonstrated that STN DBS elicits adenosine release in the striatum as measured by chemical microsensors.178 Adenosine is also known to play a role in astrocyte signaling and this fact may become even more important in view of growing interest in the local effects of DBS on glial cells.174,179,180
Glial cells far outnumber neurons in the brain and have, in recent years, been shown to play an active role in synaptic communication.181 This tripartite synapse hypothesis (involving pre- and post-synaptic neuronal elements and glia) has caused a paradigm shift in how we approach the study of neurotransmitter release and action on neural network function (see review by Perea and Araque182). There is now evidence to suggest that DBS activates glial cells directly to elicit release of gliotransmitters that, in turn, have widespread effects on the tripartite synapse and the neuronal network. As opposed to the previously presumed local inhibitory action of DBS at the site of stimulation, the prevailing effect appears to be excitation of both glial and neuronal elements and subsequent changes in neural network activity. Despite growing acceptance of this general scheme, the underlying questions of what elements are affected, how they are affected, and which neurotransmitters mediate these changes, remain largely unanswered.
It is well-documented that HFS modulates astrocyte activity by triggering the onset of a propagating Ca2+ wave.183-185 In fact, initial studies of astrocyte-neuronal interactions used electrical stimulation to evoke long-distance Ca2+ signaling186 and it is well known that electrical stimulation of brain tissue results in glial activation to increase intracellular cytosolic Ca2+ concentrations.187 Moreover, this local activation of glia can lead to a wave of Ca2+ influx that propagates through the glial cell syncytium in the brain for distances as great as several centimeters.187,188 Astrocytic Ca2+ increases, in turn, evoke the release of gliotransmitters, including ATP/adenosine, glutamate, D-serine, and PGE2.189-191 Release of these gliotransmitters can result in excitation or inhibition of neurons as well as the modulation of synaptic transmission and synaptic plasticity.181,186,192,193
Several studies have now established that astrocytes contain vesicular glutamate stores that can be triggered to undergo exocytosis by mechanical stimulation,194 in turn activating ionotropic195 or metabotropic glutamate receptors.196 In a similar manner, HFS of hippocampal slices or astrocyte cultures can elicit astrocytic Ca2+ waves.193,197 Astrocytes also release glutamate through volume-sensitive channels after ATP stimulation.198 This implies that specific stimuli may recruit different gliotransmitter release mechanisms to modify the spatio-temporal characteristics of subsequent neuronal responses. These and other studies have pointed to glutamate and adenosine as key mediators of astrocyte-to-neuron signaling.199 Astrocytes have also been implicated in mediating heterosynaptic depression, through the release of ATP and its subsequent catabolism to adenosine.174,200-202 Most notably, astrocytes respond to neuronal activity with waves of Ca2+ influx,203,204 which in turn elicit further glutamate release.205 Thus, DBS-induced Ca2+ signaling in astrocytes may affect neuronal network activity through gliotransmitter release, thereby playing an important role in the therapeutic mechanism of DBS.
DBS applied to the area containing tremor cells leads to immediate tremor arrest, an effect that is rapidly reversed when stimulation ceases.206 Similarly, we and others have demonstrated that DBS of the thalamus207,208 and STN209 results in neurotransmitter release, including glutamate. In our experimental condition, HFS applied to the thalamus led to immediate glutamate and adenosine release, which decreased to pre-stimulation levels when stimulation ceased.180 Thus, HFS-mediated glutamate and adenosine release may be important in the ability of DBS to abolish synchronized neural network oscillations such as those seen in tremor and seizures. Importantly, Bekar et al.174 have shown that thalamic DBS is associated with a marked increase in the local efflux of ATP and extracellular accumulation of its catabolic product, adenosine, which can act as a neuromodulator. Together, these findings suggest that DBS may activate multiple neurotransmitter systems, including glutamate and adenosine. Which of these neurotransmitter systems is primarily responsible for the effects of DBS in human patients and whether they are glial or neuronal in origin requires further investigation.
We and others contend that characterization of the hitherto understudied glial effects of DBS on neurotransmission will provide a deeper understanding of its corrective actions on dysfunctional brain processing, and consequently enhance our capacity to utilize its therapeutic effects in the patient.174,180 This basic knowledge will substantially enhance our potential to further develop DBS technology and surgical procedures to produce significant improvement in patient outcome. In particular, an integrative approach will be important in defining the causal relationships between DBS-mediated glial activation and neural network activity within the thalamus.
New Directions in DBS Therapy
To address the scientific and clinical issues outlined so far, it is clear that more sophisticated brain imaging and real-time in vivo electrophysiological and neurochemical monitoring techniques will be essential to mechanistic studies of DBS action, optimizing electrode placement during surgery and in the future development of intelligent, closed-loop DBS systems. As such, we will now explore these areas in turn and outline their potential roles in the evolution of DBS therapy in the future.
Optimizing electrode placement
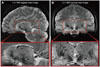
Recent advances have dramatically increased the magnetic field strength for MRI, and prototype research systems have been fitted with 7.0 T magnets, which are five-times the strength of the typical 1.5 T machines currently used in most hospitals. High-contrast and high-resolution brain images can thus be obtained with an ultra high-field MRI system to provide details never before observed in the living human brain.210-212 Fig. 1 shows sagittal and coronal images collected by 7.0 T MRI. The detailed brain structures, in particular, the high contrast visualization of the STN, should improve the accuracy of determining stereotactic coordinates for positioning stimulation electrodes during DBS surgery.213
In addition to imaging, counter-localization of the target electrode site is currently performed using microelectrode electrophysiological recordings, but in the future this may be optimized by real-time neurochemical monitoring modalities (discussed below) together with analysis of local electrophysiological parameters. In terms of the latter, recently Zaidel et al.214 in a study of 128 PD patients have shown that optimal clinical outcome of bilateral STN DBS is correlated with placement of the electrode within a distinct dorsolateral oscillatory region of the STN (characterized by increased β-oscillatory activity on multi-unit recordings), rather than simply at its anatomical centre.
Importantly, another group have shown the benefits of subsequently tailoring stimulation parameters according to the final position of the DBS electrode. In the studies by Paek et al.215 and Lee et al.216 pre-operative 1.5 T MRI images were fused with either post-operative MRI (at 3 and 6 months) and post-operative CT (at 6 months) scans respectively and the three-dimensional anatomical location of electrode contacts used to re-program stimulation parameters. In both studies, there was a significant improvement in UPDRS-III motor subscale (in both on- and off-medication states) and reduction in medication requirements.
Finally, measuring the latency of saccadic eye movements in the conscious patient (saccadometry) may also facilitate optimal electrode targeting. Temel et al.217 have shown that saccadic latency is a sensitive and objective measure of the therapeutic effect of STN DBS and it correlates well with the UPDRS-III. In the future, this could form the basis of another quantitative mode of intra-operative guidance for electrode placement in the STN.
Wireless monitoring of in vivo neurotransmitter release
Microdialysis and voltammetry are the two most widely used techniques for neurochemical monitoring in vivo.218,219 However, recording from the basal ganglia complex during clinically-relevant DBS of the STN requires a degree of temporal and spatial sampling analysis that voltammetric techniques easily provide over conventional microdialysis procedures (i.e., subsecond time scales and µm-sized space domains220). For our own aims of examining the functional anatomical and neurochemical effects of STN DBS, we developed the Wireless Instantaneous Neurotransmitter Concentration System (WINCS)-a device designed specifically to monitor neurochemical release during experimental and clinical DBS surgery. As such, research subject safety, signal fidelity, and integration with existing DBS surgical procedures, and now MRI pre-, intra, and postoperative analysis, have been key priorities during the development of WINCS. Additionally, it is easily attached to the stereotactic frame and transmits neurochemical release data to a remotely located base station (within 10 m), thus facilitating recording parameter settings and data acquisition, while minimizing encumbrance of personnel and equipment at the operating/recording site.
The WINCS device, designed in compliance with FDA-recognized consensus standards for medical electrical device safety, consists of a relatively small, wireless, sterilizable battery-powered unit that can interface with carbon-fiber microelectrodes (CFM) or enzyme-based microsensors for real-time monitoring of neurotransmitter release in mammalian brain.221-223 Indeed, our own work in pigs has utilized fast scan cyclic voltammetry (FSCV) and CFMs to evaluate neurotransmission within the basal ganglia during STN DBS. Briefly, FSCV employs a linearly scanned potential (V) applied at 100 ms intervals to brain-implanted CFMs and compared to an Ag/AgCl reference electrode in contact with the cortical surface of the brain. This detection scheme generates a voltammogram (a plot of measured oxidation and reduction current versus the applied potential) that provides a signature to identify the recorded chemical (Fig. 2). Changes in the amplitude of the oxidation peak recorded by FSCV thus provide a quantitative concentration measurement of the temporal effects of electrical stimulation on neurochemical release. Neurotransmitters and neuromodulators amenable to detection by FSCV include dopamine, serotonin, norepinephrine, adenosine, and nitric oxide.157,178,220,224-229
For our DBS studies, the WINCS device has significant advantages over other commercially available wireless recording systems as it offers 1) an advanced microprocessor with superior analog to digital conversion, greater internal memory, and faster clock speed, 2) wirelessly programmable waveform parameters (scan bias, range and rate) using an advanced Bluetooth® module for wireless communication, 3) a higher precision voltage reference for the micro-processor, 4) a low-power mode to preserve battery life, voltage sensing, and low-power alert, and most importantly 5) proven compatibility and functionality in the bore of an MRI during image acquisition. The engineered compatibility of WINCS with MRI offers a unique opportunity to quantify regional variations in neurotransmission by FSCV during fMRI procedures opening up exciting and novel research directions that go well beyond investigations of central DBS mechanisms.
Intelligent closed-loop DBS devices
DBS in its current open-loop form is always on and not guided by any changes in underlying brain activity related to the disorder being treated. However, given that the brain communicates both electrically and chemically, it is likely that the most effective treatments for many neurological and psychiatric disorders will, in the future, involve returning both electrical firing patterns and neurotransmitter release levels back to normal.230 NeuroPace (Mountain View, CA, USA) have developed a closed-loop responsive neurostimulation (RNS®) device for the treatment of refractory partial epilepsy that uses cortical surface (subdural) or depth electrodes sited at the seizure focus to monitor for abnormal electrical activity in order to elicit a pre-programmed normalizing burst of stimulation to prevent seizure onset. Alternatively, oscillations in the electrical field immediately around (and recorded using) DBS electrodes which reflect synchronized activity of large populations of neurons, termed local field potentials, have also been proposed as feedback sources for online adjustment of DBS parameters.231,232
While the ability to undertake intra- and post-operative neurochemical monitoring will be crucial in the short-term for testing predictions of the neurotransmitter release (synaptic modulation) hypothesis of DBS, in the long-term it too may form a blueprint for a closed-loop DBS device supporting all-in-one neuromonitoring and neuromodulation. Conceivably, a neuroprosthesis supporting real-time, instantaneous neurochemical sensing and feedback-guided adjustment of stimulation to maintain therapeutic neurotransmitter levels would be superior to drug treatments for neuropsychiatric disorders that entail onset and offset effects. Indeed, we have initiated work towards the ultimate realization of a closed-loop smart DBS device utilizing this rationale. One critical component is an ultra-small, low-power integrated circuit supporting wireless neurochemical monitoring. By using very-large-scale-integration techniques in standard complementary-metal-oxide-semiconductor technology, we have been able to fabricate a wireless device, with dimensions of 2.2×2.2 mm, supporting single-channel FSCV. Indeed, this device compared favorably to a conventional hardwired system in calibration tests in vitro and for measuring electrical stimulation-evoked dopamine levels in the anesthetized rat.233
Conclusion
DBS is now a widely accepted and efficacious therapy for many conditions, but we still lack a definitive understanding of its mechanism of action. Indeed, while the nature of DBS devices make them amenable to blinded trials aimed at testing the efficacy of HFS in emerging clinical contexts, future paradigm shifts in DBS technology and the movement towards closed-loop devices undoubtedly rest on our ability to gain a clearer understanding of the mechanisms responsible for the clinical effect of DBS. Fortunately, it appears that the same technological advances demanded by basic science investigators may also turn out to be central to the design of future intelligent DBS systems used by clinicians.

 XML Download
XML Download