

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
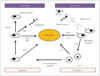
Stroke is a leading cause of disability in adults that has a heavy social burden worldwide. This disease is the third highest cause of mortality, resulting in approximately six million deaths annually.1 Acute cerebral ischemia accounts for more than 80% of all strokes and is due to brain arterial occlusion resulting from a thrombus or embolus. The pathophysiological processes following ischemic stroke are complex, involving bioenergetic failure, acidosis, loss of cell homeostasis, excitotoxicity, activation of neuronal and glial cells, and disruption of the blood-brain barrier (BBB) with infiltration of leukocytes.2 There is evidence that factors of the immune system are involved in all stages of acute cerebral ischemia (Fig. 1).3 The ischemic brain promotes a potent suppressive effect on lymphoid organs via the autonomic nervous system, which increases the risk of the poststroke infections that are major determinants of morbidity and mortality.4 On the other hand, the innate immune system contributes to subsequent repair of the damaged cerebral tissue.5
In this review we describe the main phases of the inflammatory processes during the early postischemic period, with an emphasis on the role of single mediators.
INFLAMMATION, ENDOTHELIUM, AND CLOT FORMATION
A growing amount of attention is being paid to the mechanisms of clot formation, particularly in the field of endovascular treatment of acute ischemic stroke. Although most of the focus has been on intervention devices (with developments from first- to second-generation devices, thrombus aspiration, and balloon-occlusion guiding catheters), some research groups have studied the physiopathological mechanisms of clot formation. The relation between inflammation and clot formation has been described previously, and it indicates that some cytokines (e.g., the RANTES)6 are responsible for the activation of a biochemical cascade or, indirectly, supports the concept that infections/inflammation promote atherogenesis and that some endothelial modifications that can lead to a prothrombotic status.7 Several research groups are currently focusing on the clot structure, with many findings supporting the concept of an inflammation-induced process. In particular, fibrinogen is susceptible to oxidation, and chronic exposure to oxidative stress supported by inflammation may lead to prothrombotic alterations in fibrin formation and architecture. Previous studies concerning air pollution have shown that particulate matter contributes to modulation of the fibrin structure.891011 Furthermore, some interesting studies that have used electron microscopy to investigate the clot surface (in myocardial infarction) highlight the less-investigated issue of the relation between the clot and the endothelium.12
EARLY POSTISCHEMIA TIME: THE ISCHEMIC CASCADE
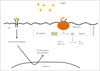
The ischemic cascade is represented by a complex series of interlinked molecular and cellular mechanisms that contribute to ischemic cell death via necrosis or apoptosis (Fig. 2). The primary insult after arterial occlusion is hypoperfusion, which dramatically reduces the availability of both glucose and oxygen in the brain, with particular vulnerability to ischemic injury being evident in specific regions: the caudate body, putamen, insular ribbon, paracentral lobule, precentral, and middle and inferior frontal gyri.13 This situation contributes to bioenergetic failure by stopping or slowing adenosine triphosphate (ATP) production.1415 A few minutes after an arterial occlusion, an ionic imbalance occurs with the abnormal influx of Na+ and efflux of K+, contributing to a widespread anoxic depolarization in the membranes of neurons and glial cells.16 The increased influx of Na+ into neurons causes the osmotic transport of water into cells that leads to cytotoxic edema, cell lysis, and necrosis. A recent neuroimaging study using 23Na MRI and quantitative histochemical K+ staining revealed heterogeneity in the rate of Na concentration increase and in the K+ distribution within the ischemic core.17 The reduced ATP production following Na/K imbalance (due to Na/K ATPase) also contributes to reduce the reuptake of glutamate, which is the main excitatory neurotransmitter.18 This condition overstimulates the glutamate receptors so as to influence the Ca2+ influx, producing a series of nuclear and cytoplasmic events (excitotoxicity) that lead to mitochondrial failure and apoptosis.1920 At the same time, the Ca2+ influx triggers the activation of catabolic enzymes with the production of arachidonic acid, and increases the formation of reactive oxygen species (ROS), mainly in neurons rather than astrocytes.21
Previous studies found that mitochondrial failure could be predicted from the K+ concentration. Indeed, the mitochondrial ATP-dependent K+ (mitoKATP) channel plays a critical role in the neuroprotective action, contributing to the so-called ischemic pre- and postconditioning states.22 The opening of mitoKATP channels attenuates the Ca2+ overload and inhibits the formation of free radicals and ROS that contribute to necrotic or apoptotic cell death.2324 The depolarization of other neurons produces a further Ca2+ influx and additional glutamate release, leading to local amplification of the ischemic damage.25 Contemporary with those processes, the persistence of arterial occlusion contributes to a critical reduction of pO2 and a concomitant increase in pCO2. In the case of hypercapnia, the tissue pH could fall to around 6.6 or lower if severe ischemia and tissue hypoxia occur; in the last situation, anaerobic glycolysis leads to lactic acid accumulation with signs of irreversible injury identifiable in the cell morphology.26 The acidosis state increases necrosis and cell death via a mechanism called acidotoxicity and mediated by Ca2+-permeable acid-sensing ion channels.272829 Other deleterious effects of acidosis influence the synthesis and degradation of cellular constituents, the mitochondrial function, the cell volume control, the postischemic flow, and the stimulation of ROS production, all conditions that occur also in the ischemic penumbra.30 Acidosis and ROS contribute to trigger a subsequent and concomitant phase represented by the activation of innate immunity and involving both resident cells (microglia) and circulating cells.
INFLAMMATORY AND INNATE IMMUNITY ACTIVATIONS
Inflammation in the ischemic brain: cell infiltration and damage
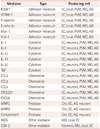
Postischemic inflammation begins in the vascular compartment immediately after arterial occlusion. The production of ROS leads to an increase in the procoagulant state involving the activation of complement, platelet, and endothelial cells.3132 The increased activity of cyclooxygenase-2 in inflammatory cells and neurons may lead to tissue damage due to excessive ROS production and toxic prostanoids.3334 ROS contribute also to reduce the availability of NO, leading to platelet aggregation and the adhesion of leukocytes, which aggravate the ischemic damage.35 There is evidence of iNOS (the inducible isoform of NO) being a critical effector and amplifier of tissue damage related to postischemic inflammation (Table 1).36 The oxidative stress and inflammatory mediators affect the permeability of the BBB, impairing the socalled neurovascular unit that includes endothelial cells, astrocytes, and neurons [involving matrix metalloproteinases (MMPs) or ROS], and allowing the extravasation of proteins and endothelial cells along with the activation of macrophages and mast cells via the ischemia and reperfusion mechanisms.37 This extravasation is also supported by a breakdown of the BBB-secondary to the pericyte death-leading to a long-lasting decrease in capillary blood flow.38
A few minutes after arterial occlusion and the associated intra- and extracellular modifications, the acute local damage is detected also by pattern-recognition receptors (PRR) (including Toll-like receptors) that respond to microbial structures (pathogen-associated molecular patterns) and host-derived danger signals (damage-associated molecular patterns).394041 These molecules can be released by stressed cells, such as during the ischemic cascade. There is new evidence that the PRR of neurons and glial cells can play a fundamental role in activating intracellular signaling pathways so as to enhance the proinflammatory expression of different genes (Fig. 3).4243 This mechanism activates the immune system elements, resulting in mast cells releasing vasoactive mediators (e.g., histamine), proteases, and tumor necrosis factor (TNF), while macrophages release proinflammatory cytokines.44
Adhesion-molecule P- and E-selectins (Table 2) and intercellular adhesion molecule-1 are expressed on the membrane after their intracellular translocation and with the rapid generation of proinflammatory signals.4546 The adhesion receptors mediate interactions between adhesion molecules and integrins, contributing to an initial rolling mechanism of leukocytes and leading to adhesion to the endothelium and to a subsequent transmigration into the brain parenchyma (leukocyte infiltration). Following ischemia, these cells rapidly release proinflammatory mediators into the area, which promotes ischemic injury via different pathways: 1) the cerebral no-reflow phenomenon by impeding the flow of red blood cells, 2) increased production of ROS and proteases at the endothelium surface, 3) platelet aggregation by leukotriene, prostaglandin, or eicosanoid production due to activation of leukocyte phospholipases, and 4) deleterious activity of proinflammatory cytokines in the penumbra area. In particular, during the acute phase of brain ischemia, microglia and macrophages release interleukin (IL)-1b, IL-6, and IL-18 from the transcriptional intracellular pathways activated by nucleotides from injured cells.47 These cytokines contribute to leukocyte infiltration in the damaged tissue, and they activate the presentation of antigens between dendritic cells and T-cells.48 T-cells lead to tissue damage by innate immunity, through interferon-gamma and ROS. T-cells activated by IL-23 released from microglia and macrophages produce IL-17, which worsens the acute ischemic cerebral injury.49 This unbalanced interplay between the immune and sympathetic nervous systems contributes to an early down-regulation of systemic cellular immune responses that leads to a functional deactivating of monocytes, T-helper cells, and invariant natural-killer T-cells.50
Previous studies have demonstrated the predictive role of certain acute immune and stress biomarkers on clinical outcomes: copeptin and mid-regional proatrial natriuretic peptide.5152 Even if the immune system is activated with recruiting elements in the focal ischemic area, specific injured cerebral sites contribute to different down-regulating responses being exhibited by the autonomous nervous system. In particular, involvement of the right frontoparietal cortex, insula, or brainstem could lead to increased cerebral inflammation and concomitant systemic immunosuppression.53 This process is characterized by the increased apoptosis of lymphocytes, suppression of peripheral cytokine release, and finally the inhibition of T-helper-1 cells and alteration of the T-helper-1/T-helper-2 ratio.54 The stroke-induced immunodepression contributes to increasing the risk of infection (influenced also by comorbidities such as age, diabetes mellitus, and atrial fibrillation) and consequently a poor functional outcome.
The activity of microglia
The microglia represent the resident immune cells of the central nervous system and can be activated by local and systemic infections, neurodegenerative conditions, and injury. Microglia are able to modify their morphology from a resting (ramified) to an active (amoeboid) state.55 Microglia respond rapidly to ischemic stroke and other injuring conditions. By entering the ischemic core within 60 minutes after the induction of focal ischemia without reperfusion, microglia significantly increase the number of their processes, while 24 hours later they are reduced in both number and distance from the insult. Previous studies have detected various forms of activated microglia.56 M1 is the proinflammatory type and is able to release TNF-a, IL-1b, IL-18, and IL-6. On the other hand, the M2 type is the healing cell involved in neuroprotection and repair, producing transforming growth factor (TGF)-b, nerve growth factor, and IL-4.57 The M1 type is observed for the first 24 hours in the ischemic core, and it increases in number over the first 2 weeks after the ischemic injury.58 The M2 type has been found at the end of the first 24 hours, entering the area during the first week before declining in number.55
Recent studies have provided evidence of microglia activation in the penumbra area. Indeed, the pattern of activity differs between the peri-infarct zone microglia and the ischemic core. During the first week, the M2 type was found to predominate in the ischemic core while the M1 type predominated in the peri-infarct zone.59 These observations suggest that the peri-infarct region is dominated by proinflammatory and activated microglia whose abundance increases during the first days after ischemic injury.60 The spatial distribution of the microglia phenotypes changes over time, which suggests the enlargement of injured and damaged cerebral tissue.
The role of inflammasomes
The above-mentioned inflammatory response leads to the production of proinflammatory cytokines and neuronal and glial cell death mediated by large intracellular multiprotein complexes called inflammasomes.45 Recent studies have demonstrated that the nod-like receptor pyrin (NLRP) and NLRP3 inflammasomes in neurons and glial cells allow detection of cellular damage and mediation of the inflammatory responses to aseptic tissue injury during cerebral ischemia.42 An increased activity of inflammasomes is also associated with neuron and glial cell death. Experimental studies and human data suggest that the NLRP1 and NLRP3 inflammasomes in brain cells could activate pro-caspase-1 (cleaving to caspase-1) to produce IL-1b and IL-18, which are proinflammatory cytokines, and lead to a particular type of cell death called pyroptosis (Fig. 3).61 In this way, the inflammasomes contribute to activating and supporting the innate immunity but also to worsening the tissue injury.
RESOLUTION OF INFLAMMATION AND REPAIR OF TISSUE
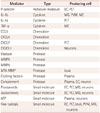
The inflammatory postischemia process is self-limiting, and its resolution is mediated by numerous factors that suppress the immune activity (Table 3). The termination of inflammation triggers structural and functional reorganization of the injured brain. The first mechanism involved in this phase is the removal of dead cells, which is performed by microglia and infiltrating macrophages, mainly comprising phagocytes.5962 The principal factors driving these cells to the ischemic site are purines released from injured cells and chemokines. Immunoglobulins directed against antigens of the central nervous system may also promote the release of IL-10 and TGF-b, which contribute to suppressing the immune process and to inhibiting the expression of adhesion molecules and the production of proinflammatory cytokines (Fig. 1).63 These pleiotropic immunoregulatory cytokines can facilitate tissue repair after promoting the resolution of inflammation, and they exert cytoprotective effects on the surviving cells in the ischemic area.35 Concomitant growth factors released by inflammatory cells, neurons, and astrocytes64 support cell sprouting, neurogenesis, and angiogenesis as well as matrix reorganization after ischemic injury. Insulin-like growth factor-1 is a critical factor in the sprouting of neurons after cerebral ischemia, while the reactivity of astrocytes is mandatory for the functional recovery of damaged tissue.65 Concomitant actions of vascular endothelial growth factor and neutrophil MMPs are required in angiogenesis, supporting the need for the combined activity of inflammatory cells and astrocytes.66
NEURONAL PRECONDITIONING AND INFLAMMATORY MEDIATORS
The brain is the most metabolically active organ in the body, consuming about 25% of total glucose and oxygen, and so a high oxidative stress is generated by its metabolism. Neurons would therefore be expected to respond actively to stress, have low capacities for replacement, and strong potential strategies for protection and repair. While most of the underlying neuroprotective and repairing pathways are still under investigation, this adaptive response to an insult by the activating intracellular signals is well known, and called preconditioning.67 Preconditioning occurs in two distinct phases. The first happens early and is associated with posttranslational modifications of proteins. This period lasts from minutes to 1–2 hours, and involves certain protective proteins (mainly protein chaperones) being rapidly released from stressed cells. The second phase requires the synthesis of new proteins. During this phase, which lasts several days, the proinflammatory cytokines IL-1b and TNF activate intracellular signaling pathways that lead to a tolerant state by mediating the stress responses. In particular, ROS and cytokines are able to activate Ras, Raf, and kinases of the cellular membrane, with the subsequent intracellular transcription and production of survival proteins. These last mechanisms contribute to preserving the endothelium via vascular protection and angiogenesis [mitogen-activated protein kinase p38 and extracellular signal-regulated kinase 1/2 (ERK 1/2)], the neurons via neuroprotection and neurogenesis [ERK 1/2 and nuclear factor kB (NF-kB)], and the glia via an anti-inflammatory action (serine/threonine kinase).68 Other proteins, called heat-shock proteins (Hsp), contribute to cellular maintenance.69 In particular, Hsp70 leads to neuronal survival, mitochondrial stabilization, and cell-death blocking, mainly by facilitating the activation of transcription factor and NF-kB.7071 NF-kB is affected also by the phosphorylation of Akt secondary to the activation of phosphoinositide-3-kinase72 This enzyme is also able to attenuate cellular apoptosis in the neurovascular unit, but it is inhibited after ischemia or reperfusion so as to induce cell death.73 Several studies have indicated that any stimulus that modifies brain function appears to increase the resistance of the brain to further injuries, including to different types of injury.74 The results of some studies involving patients with previous transient ischemic attacks suggest that brain preconditioning and tolerance occur because these patients have a more favorable clinical outcome and smaller infarcts.7576
CONCLUSIONS
Inflammatory responses during the acute stage of ischemic stroke have effects ranging from deleterious to beneficial. A first reaction is negative due to cytotoxicity, which leads to tissue damage by cellular death. At the same time, inflammation exerts protective effects by stimulating a preconditioning status that preserves brain tissue by adapting the brain itself after an insult. Finally, inflammatory mediators contribute to autolimit the pathological process and to repair the injured parenchyma by remodeling the tissue.
Future observations will contribute to a better understanding of other inflammatory mechanisms involved in these early stages of ischemic stroke, and this information will potentially lead to the development of effective neuroprotective agents.
SEARCH STRATEGY AND SELECTION CRITERIA
References for use in this review were identified by searching the PubMed and EMBASE databases for articles published between 1976 and June 2016, as well as references in relevant articles. The search terms used were “inflammation,” “immune system,” “immunity,” “brain ischemia,” “cerebral ischemia,” “stroke,” “cerebral occlusion,” and “cerebrovascular disease.” There were no language restrictions. The final reference list was generated based on the relevance to the topics covered in this review.

 XML Download
XML Download