

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Dear Editor:
Thymic stromal lymphopoietin (TSLP), a key epithelial cell and keratinocyte-derived cytokine, has been shown to directly trigger allergic inflammation and the atopic march12. Therefore, suppression of TSLP expression should be a rational therapeutic strategy for allergic disorders such as atopic dermatitis (AD). In addition, topical treatments seem to be suitable for this strategy, since TSLP is produced in epithelia such as the epidermis. Topical glucocorticoid (GC) is the most popular treatment for AD and therefore, it is rational to examine the effects of GC on the expression of TSLP in keratinocytes. In fact, it has been shown that a GC, such as dexamethasone (Dex), but not calcineurin inhibitors, suppresses the expression of TSLP in cultured normal human keratinocytes (NHK)3. In addition, it has been reported that the other GCs such as prednisolone (PSL), betamethasone, fluticasone propionate and clobetasol propionate showed an inhibitory effect on TSLP induction in NHK in the similar degree to Dex3. In the present study, we examined the effects of betamethasone butyrate propionate (BBP) on the expression of TSLP in NHK compared with Dex.
Culture of NHKs was performed as previously reported4. When NHK reached 70%~90% confluence, expression of TSLP was induced by stimulation with poly I:C (10 µg/ml; GE Healthcare), which is a toll-like receptor 3 ligand and mimics viral double-stranded RNA, tumor necrosis factor (TNF)-α (20 ng/ml; R&D Systems) and interleukin (IL)-4 (100 ng/ml; R&D Systems) with or without BBP (a gift from Torii Pharmaceutical Co., Ltd.), Dex (Sigma), or PSL (Sigma) as previously reported35. During the stimulation, we used the medium without hydrocortisone but with an equivalent concentration of ethanol (0.0001%) that was used as vehicle for GCs in all experimental groups including the negative control (i.e., absence of GCs). The NHKs were harvested 6 h after stimulation for real-time polymerase chain reaction (PCR) analysis. The supernatants were harvested 24 h after stimulation for ELISA. Isolation of total RNA, reverse transcription, and real-time PCR for analysis of TSLP expression were performed as previously reported6. The primers used for real-time PCR were the following: upper primer for human TSLP is 5′-CAG GCT ATT CGG AAA CTC AGA T-3′; lower primer for human TSLP is 5′-GTA ATT GTG ACA CTT GTT CCA GAC-3′; upper primer for human glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is 5′-TGA ACG GGA AGC TCA CTG G-3′; and lower primer for human GAPDH is 5′-TCC ACC ACC CTG TTG CTG TA-3′. Relative gene expression was calculated using the comparative Ct method or from a standard curve included in each run. Relative mRNA expression levels were normalized with a housekeeping gene, GAPDH. The quantity of TSLP in cell culture supernatants was determined by Quantikine ELISA for TSLP (R&D Systems) according to the manufacturer's instruction. All experiments were analyzed by two-tailed Student's t-test. All results are presented as means±SEMs of N experiments. A p-value of less than 0.05 was considered statistically significant.
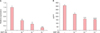
BBP suppressed the TSLP expression induced by poly I:C, TNF-α, and IL-4 at both mRNA and protein levels in a dose-dependent manner (Fig. 1). BBP reduced the expression of TSLP more strongly than either Dex or PSL at the same dose which was lower than that in the previous report3, suggesting that the suppressive effect of BBP was stronger than that of either Dex or PSL (Fig. 2).
The present study revealed that BBP not only has potent anti-inflammatory effects7 but also has a potent inhibitory effect on induction of TSLP in keratinocytes. Therefore, topical treatment with BBP might also prevent the advancement of allergic states, namely the atopic march, via direct suppression of TSLP induction in keratinocytes, although such a preventive effect of topical treatment with GCs has been confirmed in neither animal models nor atopic patients. In addition, the present study showed not only that a different GC could have a different inhibitory potential on the induction of TSLP in NHK but also that BBP might be more beneficial than other GCs, although the mechanism for this remains unclear.

 XML Download
XML Download