

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Stress has long been suspected as one of the causes of hair loss but sometimes the evidence supporting an association has been weak1. However, stress has been reported to play an important role in the pathophysiology of alopecia2345. Arck et al.6 reported for the first time that stress leads to the premature onset of catagen in mice, an effect which appears to be the result of substance P (SP) stimulation of mast cell degranulation in the context of the brain-hair follicle axis. Mast cells are regarded as the main cell type involved in stress-induced neurogenic inflammation78.
In addition to the SP-mast cell pathway, stress is known to activate the hypothalamic pituitary adrenal (HPA) axis, being the hair follicle one of the various targets of HPA axis activity910. Mouse bulge stem cells express glucocorticoid receptors, and are sensitive to the anti-proliferative effect of glucocorticoids which induce premature onset of catagen and hair growth inhibition11. We previously reported that glucocorticoids inhibit human hair growth by inhibiting both dermal papilla cells (DPCs) proliferation and growth factor expression on these cells12. In this study, we investigated the effect of acute stress on hair growth in vitro on rat and human cells, and in vivo using rat skin biopsies in the context of both, the SP-mast cell pathway and HPA axis activity.
MATERIALS AND METHODS
Acute stress animal model
All experiments involving rats were performed in accordance with the regulations and the approval of the Institutional Animal Care and Use Committee (IACUC) of the Seoul National University Hospital (IRB no. H-1102-055-351, IACUC no. 15-0023). Twenty male Sprague-Dawley rats were assigned to the experimental or control groups (ten animals per group). In the experimental group, immobilization was exerted for 24 hours using an individual rodent restrainer device (Model 544-RR; Plas Labs Inc., Lansing, MI, USA) (Fig. 1)13, which allowed rodent immobilization in a non-traumatic manner. Animals from the control group, receiving no stress, were isolated for the same time, but not immobilized. At the end of the isolation or immobilization, all animals were anaesthetized with sodium pentobarbital (50 mg/kg, intraperitoneal injection), and 4~5 ml of blood were immediately collected via intracardiac puncture, followed by cardiac perfusion with a saline solution (NaCl 0.9%, 200~250 ml per rat). Blood samples were centrifuged at 3,800g for 10 minutes, and the serum obtained was stored at −70℃.
DPC culture and proliferation assay
The methods used for isolating and culturing human DPCs (hDPCs) have been described previously14. Dr. Wendy Filsell (Unilever Research, Sharnbrook, UK) kindly provided the rat vibrissae DPC lines. Cell line culture and DPC proliferation assays were performed as described previously1516. Cell viability was determined using an MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay. All samples were run in triplicate in each independent experiment.
Analysis of cell cycle distribution
After a 72-hour serum treatment, hDPCs were fixed with 70% ethanol and incubated for 1 hour at 4℃. Cells were stained with propidium iodide (50 mg/ml) and supplemented with RNase A (Sigma-Aldrich, Steinheim, Germany) for 30 min at 4℃ prior to flow cytometry analysis. All samples were independently analyzed and repeated three times.
Stress hormone level assays
Cortisol levels and corticotropin-releasing hormone (CRH) in serum samples were measured using the commercial Cortisol EIA kit (Cayman Chemical Company, Ann Arbor, MI, USA), and the CRH Enzyme-linked Immunosorbent Assay Kit (USCN Life Science Inc., Wuhan, China), respectively, following the manufacturer's protocols.
Mast cell histology and imunofluorescence
Skin sections were deparaffinized and incubated in a 1% Alcian blue solution for 30 min. After rinsing, the sections were counterstained in nuclear fast red solution for 3 min, followed by washing. The sections were then dehydrated with 95% absolute alcohol and cleared with a xylene substitute prior to mounting. Alcian blue-positive mast cells were counted in different fields at ×100 magnification and the mean cell number per skin section was calculated.
For immunofluorescence staining, 4 µm sections were obtained from frozen skin sections and fixed with acetone at −20℃ for 15 min. Sections were blocked with blocking solution for 30 min (Golden Bridge International Labs, Mukilteo, WA, USA) and incubated overnight with an anti-mast cell tryptase antibody (Epitomics, Burlingame, CA, USA) in a 4℃ humidified chamber. The sections were then incubated with an Alexa Fluor 488-labeled goat anti-rabbit secondary antibody (Invitrogen, Tokyo, Japan), for 1 hour at room temperature. The slides were analyzed using a confocal laser-scanning microscope (Carl Zeiss MicroImaging, Inc., Thornwood, NY, USA).
Quantitative real time polymerase chain reaction and Western blot analysis
Skin samples were immediately placed into liquid nitrogen and stored at −70℃ until they were required for experiments. Quantitative real time polymerase chain reaction (qPCR) was performed as described previously using GAPDH, interleukin (IL)-1β, IL-6, IL-13, tumor necrosis factor (TNF)-α primers for rat genes17. Western blot analysis was performed as described previously using anti-β-actin, anti-mast cell tryptase monoclonal antibodies18. Samples from three independent experiments run in triplicate were analyzed.
Statistical analysis
The Mann-Whitney test was used for the hormone level assay, mast cell counts, and qPCR. The Wilcoxon Signed Rank test was used for MTT assay and flow cytometry. Results are presented as means±standard error in graphs. All quoted p-values are two-tailed, and significance was accepted at values <0.05. Data were analyzed with GraphPad Prism 6 (GraphPad Software, Inc., San Diego, CA, USA).
RESULTS
Stress serum treatment reduced the number of viable DPCs significantly
The viability of rat DPCs treated with 25% serum from stressed rats (stress serum) was lower than that of DPCs treated with 25% serum from control rats (control serum) after a 24, 48, 72-hour incubation. This effect was more apparent at 72 hours when the number of viable rat DPCs decreased significantly in stress serum-treated cells compared with those in control serum-treated cells (p<0.05, Fig. 2A). The stress serum had similar effects on hDPCs (Fig. 2B). After a 48 and 72-hour incubation, the number of viable hDPCs was significantly lesser in stress serum-treated than in control serum-treated cells (p<0.05, respectively).
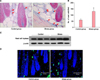
Acute stress increased the number of mast cells around hair follicles and tryptase expression
The stress induced by the 24-hour restraint increased significantly the number of mast cells in the rat perifollicular tissue. The average number of mast cells in a ×100 magnification field was significantly higher in the stress group than in the control group (n=5 per group, p<0.05, Fig. 3A, B). An increase in the skin tryptase levels in the stress group was revealed by Western blot analysis (n=3 per group, Fig. 3C). Mast cell tryptase expression was higher in the stress group than in the control group, as shown by tryptase immunofluorescence staining (n=3 of each group, Fig. 3D). Tryptase mean fluorescence intensity was 205.3%±50.9% in the stress group compared with that in the control group.
Rat skin IL-6 and IL-1β were up-regulated after restraint
qPCR showed that IL-6 and IL-1β mRNA expression levels were significantly higher in the stress group than in the control group (p<0.05, respectively). Although IL-13 and TNF-α expression levels were also higher, these increases were not statistically significant (Fig. 4).
DISCUSSION
Liu et al.19 reported that chronic stress caused by restraint inhibits hair growth via SP in mice. However, whether acute stress causes a similar inhibitory effect on hair growth remains uncertain. Based on results from our previous study showing that dexamethasone inhibits human hair growth, we hypothesized whether increased cortisol levels found in stress serum would also inhibit hair growth12. Our results confirmed that cortisol levels increased approximately two-fold after a 24-hour restraint compared with unrestrained rats. As CRH levels remained unchanged, we speculate that increased cortisol levels exerted a negative feedback on the HPA axis, leading to CRH suppression20.
The important role of mast cells in stress-induced hair loss is well known. Our results confirmed that mast cells were activated after acute stress, as demonstrated previously8. In addition, the significant increase in the total number of perifollicular mast cells we observed, may be the result of increased recruitment of mast cell progenitors in the circulation caused by restraint-induced acute stress. Mast cells originating from hematopoietic stem cells, can infiltrate the inflamed tissue within 24 hours2122. As glucocorticoid suppresses mast cell activation, such an increase in mast cell activation and numbers appears paradoxical in view of the increased cortisol levels that we found. In fact, glucocorticoids are widely used to treat allergic diseases in which mast cells are involved23. However, a suggested explanation is that acute stress may induce CRH secretion, and CRH is known to induce mast cell degranulation and increase mast cells numbers24. Although our results did not show an increase in CRH levels after a 24-hour restraint, it is possible that their levels raised earlier, triggering the cortisol negative feedback on the HPA axis. An alternative explanation involves the SP-mast cell pathway. As SP action upon mast cells is opposed to that of glucocorticoids, an increased in mast cell activation and infiltration might suggest that SP exerts a more powerful effect on mast cells than glucocorticoids, as a result of acute stress events.
SP induces the release of mast cell cytokines with inflammatory, anti-proliferative, and pro-apoptotic function which have an effect on the hair follicle epithelium25. We observed a significantly increased expression of IL-6 and IL-1β mRNA in the skin of the stressed rats. IL-6 is an important proinflammatory mediator produced by mast cells, which inhibits hair shaft elongation and suppresses proliferation of matrix cells in cultured human hair follicles2627. IL-1β is also known to inhibit hair follicle growth, and is highy expressed in patients with active alopecia areata28. Interestingly, both inflammatory cytokines, IL-6 and IL-1β are suppressed by cortisol29. However, in our study, cortisol did not fully suppress both cytokines after rats were exposed to acute stress.
Stress responses stimulate the SP-mast cell pathway as well as cortisol production via the HPA axis. The SP-mast cell pathway leads to various inflammatory reactions, and the cortisol, produced as a result of HPA axis activity, modulates such inflammation. Although the body's homeostatic mechanisms play an essential role in balancing the effects of different pathways, the SP-mast cell and HPA axis appear to exert similar effects on hair growth inhibition. Furthermore, the SP-mast cell pathway is known to play a role in the inhibition of hair follicle epithelial proliferation, and we found that serum, which contains increased cortisol after acute stress, inhibited mesenchyma-derived DPCs proliferation. These results suggest that in addition to the SP-mast cell pathway, acute stress can affect hair growth via elevated serum cortisol levels. This study has limitation that did not investigate other stress related factors besides cortisol in the serum after acute stress. Therefore it can be questioned whether cortisol was the most critical component in the serum to inhibit DPCs proliferation. Further studies to clarify more the link between acute stress and alopecia are needed. In conclusion, as well as conventional medical treatments, physicians should consider stress-reducing strategies as part of a comprehensive approach when treating patients with alopecia.



 XML Download
XML Download