

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The hair follicle (HF) is a unique mini organ in the skin of mammals that produces hair. Hair functions to thermoregulate and protect the skin, helps detect senses, and a marker for sexual development. HFs undergo a continuous cycle consisting of growth (anagen), regression (catagen), and relative quiescence (telogen) phases. Growth of new hair shafts and regression of old hair shafts are repeated through this cycle1. Multiple signaling pathways and genes are involved in regulating the hair cycle and HF morphogenesis2-4.
Wnts are a family of secreted glycoproteins that play a critical role in the embryonic development and maintenance of homeostasis in mature tissues by regulating proliferation, differentiation, migration, and apoptosis of cells5-8. The Wnt pathway regulates the regeneration and differentiation of HFs9-11. Wnt signaling pathway activity is regulated by secreted inhibitors, which are divided into two functional classes. One class includes the secreted frizzled-related protein (Sfrp) family, Wnt inhibitory factor 1 (Wif1), and Cerberus which binds directly to Wnt, thus, sequestering Wnt from its receptors. The Dickkopf (Dkk) class binds to the LRP5/LRP6 component, thereby inhibiting binding of Wnt to its receptor12.
The other class, sFRP, a family of glycoproteins, has five members (Sfrp1~Sfrp5) that participate in the developmental and disease processes in various cells and tissues by controlling the Wnt pathway13,14. The sFRP family possesses structural similarities to the cysteine-rich domain of frizzled receptors; thus, they interact with Wnt and form the Wnt-Sfrp complex. Therefore, the Wnt protein is sequestered from the frizzled receptor15.
Sfrp2 has been studied in different cell types and organs to determine its role16-18. Although Sfrp2 is expressed in the upper and lower matrix, outer root sheath, and dermal papilla19,20, its role in HFs has not been investigated.
Thus, we investigated Sfrp2 expression at various HF stages in mouse dorsal skin and the effect of SFRP2 on keratinocytes to identify the role of SFRP2 in HFs. We found that Sfrp2 expression peaked at the catagen phase, and that this expression pattern was inversely related to that of Wnt target genes. We also observed inhibited mouse keratinocyte proliferation by SFRP2. These results suggest that Sfrp2 may play an important role during the catagen phase by inhibiting keratinocyte proliferation in HFs.
MATERIALS AND METHODS
Animal
BALB/C mice were purchased from Orient Bio (Seongnam, Korea) and maintained in the barrier system with regulated light (700 to 1,900 h), temperature (23±1℃), humidity (50%±5%), and ventilation (10 to 12 times/h). All animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of the Catholic University of Korea and were carried out in accordance with the Guidelines for Animal Experimentation.
Histological study
Mouse dorsal skins of BALB/C mice at postnatal days 10 (P10), P14, P17, P21 and P28 were gathered as previously described21. All skin samples were harvested from the same region (2×2 cm) of the upper back skin. Six-micrometer-thick paraffin sections were prepared. Hematoxylin and eosin (H&E) staining was carried out using the standard method, and the stained sections were observed with an optical microscope (Olympus, Tokyo, Japan).
RNA isolation and reverse transcription
Total RNA was extracted from the dorsal skins of BALB/C mice at P10, P12, P14, P17, P21, P28, and P35 using TRIZOL following the manufacturer's instructions (Invitrogen, Camarillo, CA, USA). Two micrograms of RNA were utilized to synthesize single stranded cDNAs using the PrimeScript 1st strand cDNA Synthesis kit following the manufacturer's instructions (TaKaRa Bio Inc., Shiga, Japan).
Reverse transcription-polymerase chain reaction and Real-time polymerase chain reaction
Polymerase chain reaction (PCR) was performed using Thermal Cycler-100 (MJ Research Inc., Waltham, MA, USA). PCR conditions were as follows: initial denaturation for 2 minutes at 95℃ followed by 28~30 cycles of 15 seconds at 94℃, 15 seconds at 62℃, and 15 seconds at 72℃. The final extension was performed for 10 minutes at 72℃. Real-time PCR was performed with the cDNAs prepared from the total RNAs of the skin at various time points (See the above section) using SYBR Premix Ex Taq (TaKaRa Bio Inc.) in an Mx3000P (Stratagene, La Jolla, CA, USA). Specific primer sequences for each gene are listed in Table 1. The cycling conditions were as follows; initial heating for 2 minutes at 95℃ followed by 45 cycles of 15 seconds at 94℃, 15 seconds at 62℃, and 15 seconds at 72℃, and the final extension was performed for 10 minutes at 72℃. Gene expression levels were determined by the comparative ΔΔCt method using the expression level of Gapdh as control22. Relative mRNA expression levels were determined based on Real-time PCR performed in duplicate using various numbers of independent samples for each point (number of sample for each point; P10=3, P12=4, P14=7, P17=4, P21=5, P28=5, P35=4).
Plasmids
The 888 bp of Sfrp2 coding sequence (CDS) were amplified by PCR using Expand High Fidelity enzyme (Roche Diagnostics, Basel, Switzerland) from skin cDNAs of BALB/C mice. Forward and reverse primer sequences are listed in Table 1. PCR products were subcloned into pcDNA 3.1 using EcoRI cloning sites (Invitrogen).
Cell culture and transfection
Mouse keratinocyte cells (PAM212 cell line) were cultured in DMEM (Invirogen) containing 10% FBS with 5% CO2 in a 37℃ incubator. Transfection experiments were performed using polyethyleneimine (Sigma-Aldrich, St. Louis, MO, USA) according to the manufacturer's instructions. A total of 8×105 cells were plated in 60 mm dishes in triplicate. Following this, either 500 ng of pcDNA 3.1/ Sfrp2 CDS or only pcDNA 3.1 plasmid was transfected into the cells with 0.4 µg of pCMV3.1/β-gal. Transfection experiments were normalized against transfection efficiency determined by β-galactosidase activity. To ensure that Sfrp2 overexpression was well induced, we performed Real-time PCR and found a 27118-fold increase in the Sfrp2 mRNA level compared with that of pcDNA 3.1 transfection. After incubation for 48 hours at 37℃, the transfected cells were observed using a microscope (Olympus) or harvested for extraction of total RNAs.
SFRP2 protein treatment
SFRP2 protein was purchased from R&D system and reconstituted in sterilized phosphate buffered saline (PBS). A total of 5×105 PAM212 cells were plated on a 100 mm culture dish in DMEM media and were treated with SFRP2 the following day by replacing media with the media containing SFRP2 (50 ng/ml or 100 ng/ml). The treatment was continued for 3 days by changing the media every day.
Immunocytochemistry
For immunocytochemistry, PAM212 cells either treated with SFRP2 or transfected with pcDNA 3.1/Sfrp2 were washed three times with cold PBS and fixed using 4% paraformaldehyde for 10 minutes at room temperature. The cells were then treated with 0.5% Triton X-100 for 10 minutes and washed three times with PBS. After blocking with 3% bovine serum albumin for 1 hour, the cells were incubated with antibodies against Ki67 (Thermo Scientific, Hudson, NH, USA; 1:200) or Involucrin (Covance, Berkeley, CA, USA; 1:200) overnight. After washing, Alexa Fluor 488 goat anti-rabbit secondary antibody was applied for 3hr (Invitrogen, 1:500), and 4 g/ml of Hoechst 33342 (Sigma-Aldrich) was also applied for 1 minute to stain the nuclei. Fluorescence signal was observed with a Fluorescent microscope (Olympus).
RESULTS
Sfrp2 mRNA expression during the hair cycle
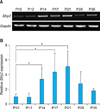
To determine the role of Sfrp2 in HFs, first we investigated the relative expression level of Sfrp2 compared to those of other Wnt inhibitors using reverse transcription-polymerase chain reaction (RT-PCR) and Real-time PCR. Among the Wnt inhibitors with relatively high expression at P10, Sfrp2 was the only one whose expression was dramatically increased at P17. Furthermore, the expression level of Sfrp2 was 5.9-fold higher than that of the second most expressed Wnt inhibitor, wif1 (Fig. 1). These results suggest that Sfrp2 is the most abundantly expressed Wnt inhibitor in HFs at the catagen phase.
Relative expression level of Sfrp2 mRNA was determined at different hair cycle stages by RT-PCR as well as Real-time PCR analyses. We found that Sfrp2 was highly expressed during catagen, which was maintained until the early telogen phase (Fig. 2A). During anagen, Sfrp2 expression was weaker than those of other stages. After this, the expression level increased significantly by 2.4~3.6 fold as the stages progressed into the catagen and early telogen phases. As the next anagen phase began, Sfrp2 expression started to decline and then decreased again to a low level that is similar to that of the first anagen phase by P35 (Fig. 2B). These Sfrp2 mRNA expression patterns suggest that Sfrp2 may function during the catagen phase.
Effect of SFRP2 on keratinocyte proliferation
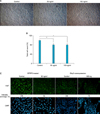
During the catagen phase, HFs undergo degeneration through cessation of proliferation and subsequent differentiation of keratinocytes. Because Sfrp2 expression increased at the catagen phase as shown in Fig. 2, we investigated whether Sfrp2 is involved in the regulation of proliferation and/or differentiation using the PAM212 mouse keratinocyte cell line. Cells were treated with SFRP2 at 50 and 100 ng/ml for 48 hours, and the number of live cells was counted using the trypan blue exclusion method. We found that SFRP2 significantly reduced the number of cells by 20% compared to the mock-treated control at both 50 and 100 ng/ml (Fig. 3A, B).
To investigate whether this decrease was caused by inhibition of cell proliferation, we used immunofluorescent staining for the Ki67 proliferation marker23. Immunocytochemistry showed decreased Ki67 expression in cells treated with SFRP2 and in cells overexpressing SFRP2 (Fig. 3C), indicating that SFRP2 inhibits keratinocyte proliferation.
Effect of SFRP2 on Involucrin expression in keratinocytes
To investigate whether SFRP2 treatment would affect the differentiation status of keratinocytes, we performed immunocytochemistry for Involucrin expression, a keratinocyte differentiation marker24, in SFRP2 treated or Sfrp2-overexpressed PAM212 cells. Unlike Ki67, we found no difference in Involucrin expression between control and SFRP2-treated cells (Fig. 4A).
RT-PCR and Real-time PCR analyses also revealed no significant difference in the Involucrin mRNA level between control and SFRP2-overexpressed cells (Fig. 4B, C). These results indicate that Sfrp2 does not affect Involucrin expression and suggest that Sfrp2 is not involved in keratinocyte differentiation.
Effect of SFRP2 on the Wnt pathway during the hair cycle
SFRP2 function in the Wnt pathway is controversial. SFRP2 was reported to be inhibitory in the Wnt pathway12, whereas it was also shown to activate Wnt signaling through the canonical activation of β-catenin in intestinal epithelial cells and canine mammary glands25-28. To determine SFRP2 function in the Wnt pathway of HFs, we investigated the expression of the wnt target genes Ccnd1 and C-myc on the back skin of mice. Real-time PCR analysis revealed an inverse relationship between the Ccnd1 and C-myc expression pattern and that of Sfrp2 throughout the HF cycle (Fig. 5), suggesting that Sfrp2 may function as a Wnt inhibitor in HFs.
DISCUSSION
Many signaling pathways crosstalk with each other to control HF development and cycling29. Among these signaling pathways, the Wnt pathway has been widely accepted to function in hair morphogenesis and HF cycling30-33. Excess induction of Wnt signaling causes abnormal hair cycles and abnormal formation of HFs34. Several mice with Wnt inhibitor mutations have been reported. For example, a Dkk1 or Dkk2 mutation in mice causes complete hair loss through failure to initiate development and mis-spacing of HFs6,9, showing that Dkk1 and Dkk2 are essential in normal HF formation and control of the hair cycle. Although mice with the Sfrp2 mutation have been reported and studied35, its function in the normal hair cycle has not been documented.
In this study, we found that Sfrp2 is mainly expressed during the catagen phase and inhibited mouse keratinocyte proliferation. This inhibition was also observed in human HaCaT keratinocytes (data not shown). These results suggest that Sfrp2 may function in the catagen phase by inhibiting keratinocyte proliferation. The inhibition rate by SFRP2 in keratinocytes was 20%, suggesting that SFRP2 may not be the main catagen regulator. Nevertheless, SFRP2 must be considered a contributor in the catagen phase of HFs.
Several investigators have shown that the action of Sfrp2 in the Wnt pathway differs in different cell types. These studies suggest that Sfrp2 acts not as a Wnt inhibitor but as an activator25,26. However, in our study, we showed that Sfrp2 expression is inversely related to Wnt target gene expression (Fig. 5), suggesting that Sfrp2 is a Wnt inhibitor in HFs. In addition to Sfrp2 expression, other Wnt inhibitors are also expressed in HFs (Fig. 1). Thus, Wnt target gene expression in HFs must be the sum of Wnt inhibitor expression. Therefore, although Sfrp2 acts as a Wnt inhibitor in the normal hair cycle, a further study is required to understand its precise function in HFs.
SFRP2 is also known to regulate the differentiation of myoblasts and osteoblasts18,36. Our study found that SFRP2 treatment on keratinocytes did not affect the expression of Involucrin, a differentiation marker of keratinocytes. Furthermore, there was no significant difference between the control and SFRP2 treated human HaCaT keratinocytes (data not shown). These results suggest that SFRP2 may not play a role in the differentiation of keratinocytes.
In conclusion, our results suggest that SFRP2 plays a role in the catagen phase by inhibiting keratinocyte proliferation. A further study with microdissection of the epidermis is needed for a better understanding of SFRP2 function in hair cycle regulation.




 XML Download
XML Download