

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Melanin plays an important role in protecting the skin from ultraviolet light. It also determines skin color and influences phenotypic appearances. However, abnormal accumulation of melanin may lead to esthetic problems. Hydroquinone (HQ) is a major ingredient in topical pharmacological agents that are used for hyperpigmentary disorders. However, HQ is frequently associated with a high rate of adverse effects1,2.
Therefore, several topical hypopigmenting agents have been developed and widely used (Table 1). This review summarizes the different approaches that have been implemented to achieve hypopigmentation and classify them on the basis of their mechanisms.
TOPICAL HYPOPIGMENTING AGENTS HQ IN MELASMA
Treatment of melasma is difficult and a number of agents have been used for this intractable condition. To date, the most effective treatment is a triple-combination cream that contains 4% HQ, 0.05% tretinoin and 0.01% fluocinolone acetonide3. HQ is the most commonly used tyrosinase inhibitor. Oxidation products of HQ result in oxidative damage of membrane lipids and proteins, including tyrosinase, and depletion of glutathione4. However, HQ is not commonly used in cosmetics because of long-term complications5. Tretinoin is used as an anti-wrinkle agent. However, it is also reported that topical retinoid is effective in the treatment of pigmentary disorders or can be combined with other topical agents6. The third ingredient featured in triple combination products is corticosteroids. Steroids are effective in the suppression of cytokines such as endothelin-1 and granulocyte macrophage colony-stimulating factor (GM-CSF), which mediate ultraviolet (UV)-induced responses7,8. By using a triple-combination cream, combined effects can be expected for the treatment of pigmentary conditions.
The efficacy and safety of triple-combination creams have been reported in several studies9. In one study, 26.1% showed a complete clearing of melasma by week 810. In another multicenter study, global evaluations revealed that 75% of patients showed "moderate or marked improvement" or were "almost clear" or "clear" by week 8. These results showed that a triple-combination cream is a rapidly effective topical agent for the treatment of melasma11. However, in one study, 57% of patients experienced at least one treatment-related adverse reaction, although it was claimed to be mild and transient in nature12. In particular, post-inflammatory hyperpigmentation should be expected in darker-skinned patients13. A multicenter study was also performed to compare the efficacy and safety of a triple-combination cream in Asian patients. The results showed that a triple-combination cream was also effective in Asian patients with tolerable adverse reactions. Although most adverse events were mild, almost 50% of patients reported the incidence of related adverse events14. Thus, it should be noted that triple-combination creams are effective for melasma, but can have a high frequency of adverse reactions.
CLASSIFICATION OF TOPICAL AGENTS FOR PIGMENTARY CONDITIONS
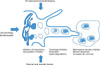
At present, the most effective hypopigmenting agents are tyrosinase inhibitors. However, melanogenesis can be controlled by regulating (i) the transcription and activity of enzymes such as tyrosinase, tyrosinase-related protein-1 (TRP-1), tyrosinase-related protein-2, and/or peroxidase; (ii) the uptake and distribution of melanosome in keratinocytes and (iii) melanin and melanosome degradation and turnover of "pigmented" keratinocytes5. However, it is very clear that hypopigmenting agents can work by different combined mechanisms. Furthermore, melanogenesis is controlled by additional factors via keratinocytes, fibroblasts and also by local and/or systemic conditions (Fig. 1).
Regulation of enzymes
1) Regulation of transcription and maturation of tyrosinase
Transcription of genes encoding tyrosinase and TRP-1 is under the control of the microphthalmia transcription factor (MITF)15. Mitf is a critical transcription factor for both melanocyte proliferation and melanogenesis. As Mitf is regulated by the Wnt signaling pathway as well as cAMP, and by both p38 signaling and the MAP kinase pathway, any agents that can potentially regulate these signaling pathways will also affect Mitf and melanogenesis. Although it is not clinically available, sustained extracellular signal regulated kinase (ERK) activation by sphingosine-1-phosphate (S1P) can lead to MITF phosphorylation and degradation, which in turn are responsible for decreased melanin synthesis16. Transforming growth factor (TGF-β1) also plays an inhibitory role in melanogenesis. TGF-β1 induced a significant delay in ERK activation and ERK-induced down-regulation of Mitf17. Furthermore, lysophosphatidic acid and C2 ceramides are able to induce Mitf degradation or decrease Mitf expression18-20.
These are examples that inhibit melanogenesis by transcriptional regulation of the tyrosinase gene. Tyrosinase is a glycosylated protein. Therefore, glucosamine or tunicamycin, which are specific inhibitors of lipid carrier-dependent glycosylation, can induce hypopigmentation21. In addition, calcium D-pantetheine-S-sulphonate (PaSSO3Ca) causes an inhibition of melanogenic enzymes possibly through the alteration of tyrosinase and TRP-1 glycosylation without affecting their expression22.
2) Inhibition of tyrosinase activity
There are several tyrosinase inhibitors that have been used to produce hypopigmenting topical agents or cosmetics. Arbutin, a naturally occurring HQ beta-D-gluconopyranoside is commonly used23. Arbutin decreases tyrosinase activity without affecting mRNA expression and inhibits 5,6-dihydroxyindole-2-carboxylic acid (DHICA) polymerase activity24. Kojic acid (5-hydroxy-2-hydroxymethyl-4H-pyran-4-one) was also a commonly used antibiotic agent produced by species of Asperigillus and Penicillum25.
Among these, 4-n-butylresorcinol has been characterized as a strong tyrosinase inhibitor26. We compared the hypopigmenting effects of HQ and 4-n-butyl resorcinol and showed that HQ (100µM) and 4-n-butyl resorcinol (10µM) had similar tyrosinase inhibition activities. These finding suggests that 4-n-butyl resorcinol is a more potent tyrosinase inhibitor than HQ. Thus, the hypopigmenting effect can increase if the concentration of 4-n-butyl resorcinol is increased. However, a cautious increase will be necessary to ensure both efficacy and safety. Positive hypopigmenting effects of 4-n-butyl resorcinol have previously been demonstrated27.
Generally, phenolic compounds are known as hypopigmenting agents because of their ability to serve as alternative substrates for tyrosinase28.
3) Post-transcriptional control of tyrosinase
There are examples of agents that inhibit melanogenesis by the increased degradation of tyrosinase proteins. Unsaturated linoleic acid decreases tyrosinase activity, whereas saturated palmitic or stearic acids increases tyrosinase activity. Also, topical application of linolenic, linoleic and oleic acids produce a hypopigmenting effect on guinea pig skin stimulated with UV light29. It is reported that linoleic acid decreases the amount of tyrosinase through increased tyrosinase ubiquitination and degradation by the proteasome30,31. In addition, other agents like phospholipase D2 decrease melanogenesis through the same ubiquitin-mediated degradation of tyrosinase32.
Inhibition of melanosome transfer
Melanosomes are specialized organelles in which melanin is synthesized and deposited. The addition of TGF-β1 to cultured melanocytes produced less pigmented melanosomes even when the cells were concomitantly treated with αMSH to increase their fully melanized melanosomes33. It is also reported that ERK activation by S1P can lead to hypopigmentation16. Interestingly, decreased melanization of melanosomes was also found in S1P treated melanocytes (unpublished data). Moreover, S1P-treated cells showed undifferentiated early stage melanosomes, whereas control cells showed internal fibrils and dense pigments in melanosomes. These findings suggest that inhibition of melanosome formation can be a good strategy to control melanogenesis. However, there are no agents that are currently available to meet this requirement.
Melanosome formation is an important step in melanognesis and melanosomes need to be transferred to keratinocytes from melanocytes for the completion of this step. Thus, the inhibition of melanosome transfer can produce a hypopigmenting effect. Theoretically, the inhibition of serine protease can result in an impaired activation of protease-activated receptor 2 on the keratinocyte leading to the accumulation of melanosomes within the melanocyte34. Clinically, niacinamide (Vitamin B3), which is commonly used to manufacture cosmetics, has been found to inhibit melanosome transfer to keratinocytes both in vitro and in vivo35.
Additional mechanisms
1) Regulation of melanocytes environment
Endothelin 1 (ET-1), which is produced by keratinocytes after exposure to inflammatory stimuli or UV exposure, stimulates melanogenesis. ET-1 has strong stimulatory effects both on DNA synthesis and melanization in human melanocytes. Thus, topical application of M. chamomilla extract inhibits ultraviolet B (UVB)-induced pigmentation by inhibiting ET-1 effects36.
Clinically, it is well-known that topical corticosteroids have strong anti-inflammatory effects. They have been used for the treatment of melasma to decrease irritation caused by hypo-pigmenting agents37, and work by the suppression of cytokines through the inhibition of nuclear factor kappa B (NF-κB) activation. Topical steroids can be effective by the suppression of cytokines such as endothelin-1 and GM-CSF, which mediate UV-induced pigmentation7,8.
There are several ingredients with anti-inflammatory activity. Glabridin, the main component of hydrophobic fraction of licorice extracts decreases tyrosinase activity in B16 melanoma cells and inhibits UVB-induced skin pigmentation as well as erythema. The capability to inhibit cyclooxygenase activity and superoxide anion production implies that this anti-inflammatory effect requires an interference with the arachidonic acid cascade. Consequently, protection against oxidative stress plays a key role in controlling melanogenesis38.
2) Antioxidant agents
In general, antioxidants exhibit hypopigmenting effects by interacting with o-quinones, thus avoiding the oxidative polymerization of melanin intermediates, or with copper at the active site of tyrosinase. In addition, antioxidant agents can regulate the signaling process by scavenging ROS in the skin39. For example, ascorbic acid can interfere with melanization by interaction with copper ions at tyrosinase and reduction of dopaquinone and DHICA oxidation40,41. α-Tocopherol and its derivatives can also regulate melanogenesis. The antioxidant property affects the lipid peroxidation of membranes and increases the intracellular glutathione content42. 6-Hydroxy-3,4-dihydrocumarins, another novel type of antioxidant, have an anti-melanogenic activity in cultured normal human melanocytes at non-cytotoxic concentrations without interfering with tyrosinase activity43. The acceleration of glutathione synthesis and the inhibition of tyrosinase transfer may be the mechanism of action44. α-lipoic acid, a disulfide derivative of octanoic acid, has been reported to prevent UV-induced oxidative damage, mainly through the down-modulation of NF-κB activation. In addition, this agent is known to inhibit tyrosinase activity by possibly chelating the copper ions45. Peroxidase is involved in the polymerization of melanogenic intermediates46. Based on this, the inhibition of peroxidase can decrease melanogenesis by reducing the polymerization of eumelanin47. Methimazole, an antithyroid agent belonging to the thionamide group, shows inhibitory action towards tyrosinase and peroxidase48. Mild to moderate inhibition of melanization can be expected with morphological changes of melanocytes in animal models.
3) Combination of multifunction hypopigmenting agents
Recently, we reported that terrein, a bioactive fungal metabolite isolated from a Penicillium species, reduces melanin synthesis by reducing tyrosinase production via ERK activation, and that this is followed by MITF downregulation49. Interestingly, we also found that terrein decreases melanogenesis through ubiquitin-dependent proteasomal degradation as well as decreased expression of its mRNA50. Thus, terrein can be an example of a hypopigmenting agent that inhibits melanogenesis by dual action including the down-regulation of transcription and up-regulation of degradation.
In contrast to terrein with multifunction, the combined use of two agents with different action mechanisms can be additive in terms of total effects. As already described, 4-n-butylresorcinol did not induce ERK or Akt activation, or MITF degradation, and also had no effect on cAMP response element binding protein phosphorylation, which stimulates MITF expression26. However, 4-n-butylresorcinol showed an additive effect in combination with hinokitiol, which reduces MITF expression. Thus, the combination of these two agents with different action mechanisms can be another strategy to increase the efficacy of these agents.
CONCLUSION
Recently, melanocyte biology has made remarkable progress. However, the pathogenic mechanisms underlying acquired hyperpigmentation have not been completely understood. Even though these mechanisms need to be explored further, our present understanding has improved greatly, which has contributed to the enhancement of diagnosis and treatment of pigmentary conditions. In particular, the finding that the dermal microenvironment can affect epidermal pigmentation through dermal degeneration or vascular dilatation has had a significant influence. These findings suggest that environmental effects on melanogenesis are very important. In addition, the combined use of multiple agents with different actions can show additive effects. As already described, 4-n-butylresorcinol is a strong tyrosinase inhibitor, whereas a signal regulator such as terrain affects pigmentation through ERK-induced MITF degradation. Thus, the combination of multiple agents with different mechanisms of action can be another strategy to increase the efficacy of these agents.

 XML Download
XML Download