

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Malassezia folliculitis, as with seborrheic dermatitis, affects sites where there is an enhanced activity of sebaceous glands such as the face, upper trunk and shoulders. These patients often present with mild pruritus or follicular rash and pustules without itching1,2. It usually occurs in the setting of immuno-suppression such as the use of steroids or other immunosuppressants, chemotherapeutic agents, bone marrow transplantation and diabetes. The etiology is presumed to be overgrowth of Malassezia yeasts, which normally exist as harmless resident flora of the skin1,2.
Malassezia yeasts, which are known to be the culprits of seborrheic dermatitis, pityriasis versicolor and neonatal pustulosis, have recently taken the center stage. Their central role as the aggravating factor of atopic dermatitis has become the center of attention for the past several years and the number of research articles and case reports on this has shown a rapid increase3-6. However, despite a high level of interest, there have been only scarce studies on the inter-relationship between Malassezia and Malassezia folliculits to this point.
In response to the growing demands, the authors of this study, with the aid of 26S rDNA PCR-RFLP, sought to find clues on this inter-relationship by showing how the frequency and distribution of Malassezia yeasts vary according to age, gender and the body site, and we also wanted to compare the results with those of a healthy control group.
MATERIALS AND METHODS
Subjects
The patient group consisted of sixty Malassezia folliculitis patients (30 males and 30 females; 20 in their teens, 20 in their twenties and 20 in their thirties) who were diagnosed with the disorder from July of 2005 to December of 2009 at Konkuk University Hospital, the outpatient Department of Dermatology, and these 60 patients were matched with sixty healthy controls. Excluded from the study were those patients who were on systemic adrenocorticoid therapy, phototherapy or antifungal agents within the past two months, as well as those who had been treated with topical antifungal agents within the past month and/or with topical corticosteroid within one week. All the subjects were instructed to avoid the use of moisturizers and facial cleansing on the day of examination, and informed consent was obtained from each individual after providing a thorough explanation of the possible physical and psychological adverse outcomes that may arise during the course of the study. This study was conducted in strict abidance with the principles of the Declaration of Helsinki.
Culture of the clinically isolated species
1) Preparation of the Leeming-Notman medium and washing solution
Leeming-Notman agar medium was prepared by mixing glycerol monoesterate (BDH, Poole, UK) 0.5 g, bacteriological peptone (Oxoid, Hampshire, UK) 20 g, glucose (Oxoid) 5 g, yeast extract (Oxoid) 0.1 g, ox bile (Merck, Darmstadt, Germany) 4 g, agar No.1 (Oxoid) 12 g, Tween 60 (Yakuri, Osaka, Japan) 0.5 ml and glycerol (Tedia, Fairfield, CA, USA) 1 ml with one liter of distilled water and then sterilizing it for twenty minutes at 121℃. Following the sterilization process, cycloheximide (Sigma, St Louis, MO, USA) 200 mg and chloramphenicol (Sigma) 50 mg were added, followed by 5 ml of non-skim milk treated at a super-high temperature (Konkuk Dairy, Seoul, Korea). After thorough mixing, the solution was spread evenly on a petri dish and kept refrigerated until use. Washing solution was prepared by dissolving NaH2PO42H2O 1.17 g into 100 ml of distilled water; 85 ml of which was taken and mixed with Na2HPO4 10.6 g and 1,000 ml of distilled water. The pH was adjusted to 7.9, and 1 ml of Triton X100 was added to the solution before sterilizing it at 121℃ for twenty minutes and then refrigerating it.
2) Specimen sampling
Specimens were harvested from forehead, cheek and chest, and the specimens were sampled by a scrub-wash technique, based on the method suggested by Williamson and Kligman7. A stainless tube with an interior area of 4.909 cm2 was set on the selected part of the skin (forehead, cheek and chest) and then 1 ml of detergent (0.01% NaH2PO42H2O, 1.01% Na2HPO4, 0.1% Triton X-100 [pH 7.9]) was added to the tube. After rubbing the skin with a glass rod for one minute, the sample was removed using a pipette and stored in a different container. Then, 1 ml of the detergent was added to the stainless tube, and the specimen was repetitively sampled and added to the first sample. One hundred microliters of the sampled specimen was then mixed with 900µl of the detergent, and 100µl was taken from the mixture, evenly applied on the Leeming-Notman medium and cultured at 34℃ for 14 days.
Molecular analysis
1) DNA extraction and PCR
For DNA extraction and PCR analysis of the skin isolates, we adopted colony PCR analysis8, which was developed for extracting the DNA directly from a colony of a PCR tube and for amplification of 26S rDNA at the same time instead of using the direct genomic DNA extraction methods. A colony of Malassezia yeast was removed, transferred to a PCR tube and warmed 3 times a day for 1 minute each time in a double boiler using a microwave; the tube was then immersed in ice water. The PCR reaction mixture (0.25 mM deoxynucleoside triphosphate, 10X PCR buffer, 5X Q buffer, 0.5µM primers, 1.25 U Hot StarTaq polymerase, 20 mM MgSO4) was added and vortex mixing was performed. Then, PCR was performed immediately using a Mastercycler 5333 (Eppendorf, Hamburg, Germany). To amplify the 26S rDNA, a primer that is capable of amplifying all 11 standard strains at once was chosen. The sequence was as follows: forward, 5'-TAACAAGGATTCCCCTAGTA-3' and reverse, 5'-ATTACGCCAGCATCCTAAG-3'. The conditions in the early stage of the reaction were 95℃ for 14 minutes for pre-denaturation, 94℃ for 45 seconds for denaturation, 55℃ for 45 seconds for annealing, 72℃ for 1 minute for extension of 40 cycles, and 72℃ for 7 minutes for the last extension. The amplified DNA was visualized by electrophoresis on 1.5% (w/v) agarose gel with ethidium bromide (0.5µg/ml) staining and using 1X TAE migrating buffer (pH 8.0, 40 mM Tri-acetate 1 mM EDTA).
2) Restriction fragment length polymorphism analysis
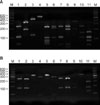
After checking the amplified 26S rDNA, the product of PCR was purified using an Accu-Prep PCR purification kit (Bioneer, Daejeon, Korea). For the 26S rDNA RFLP analysis of Malassezia yeasts, two restriction enzymes were used: Hha I (Takara Biomedicals, Otsu, Japan) and BtsF51 (SibEnzyme, Novosibirsk, Russia). The restriction enzyme reaction was conditioned with 10X PCR buffer and 10 U restriction enzyme, and the reaction mixture included 7.5µl of the PCR product. After three hours of reaction at 37℃, electrophoresis was performed in TAE buffer on 3.5% (w/v) NuSieve GTG agarose gel (FMC, Rockland, ME, USA). The gel was then stained with ethidium bromide, and the size and number of DNA fragments were measured by a UV transilluminator for analysis of the RFLP patterns (Fig. 1).
RESULTS
Comparison of the positive culture rates of the Malassezia folliculitis patients and the healthy controls
Among the 60 Malassezia folliculitis patients, Malassezia yeasts were isolated in 106 of 180 (58.9%) samples collected from the forehead, cheek and chest, and five species of Malassezia (M. restricta, M. sympodialis, M. furfur, M. globosa and M. dermatis) were identified. The same five species were also identified in the healthy controls. By age, the teens group (age 11~20) had the highest rate of detection at 63.3% (38 of 60), followed by the thirties group (37 of 60, 61.7%) and the twenties group (31 of 60, 51.7%). In the control group, the thirties group had the highest rate (45 of 60, 75%) (Table 1). According to site, the chest had the highest detection rate at 63.3% (38 of 60), followed by the forehead (37 of 60, 61.7%) and the cheek (31 of 60, 51.7%). In the control group, the rate was highest for the forehead (51 of 60, 85%) (Table 2).
Species of Malassezia yeasts by age
By age, M. restricta was the most frequently isolated species in the teens (23.3%) and twenties groups (25.0%), while M. sympodialis was the most predominant species in the thirties group (30%). In contrast, in the control group, M. globosa was the most predominant species in the teens (28.3%) and twenties (33.3%) groups, while M. sympodialis was most commonly isolated in the thirties group (25%) (Fig. 2). Statistical significance was detected in the teens and twenties groups, but not in the thirties group (Table 3).
Species of Malassezia yeasts by the body site
Overall, M. restricta was the most frequently isolated species (37 of 180, 20.6%). On the forehead, M. restricta was found in 28.3% of the cases, M. sympodialis was found in 20% and M. globosa was found in 6.7%. On the cheek, M. restricta was isolated in 20% of the cases, M. sympodialis was isolated in 16.7% and M. globosa was isolated in 10.0%. On the chest, M. restricta was isolated in 13.3%, M. globosa was isolated in 21.7% and M. sympodialis was isolated in 18.3%, while M. furfur was found at a significantly lower frequency, with a 10% detection rate. In the control group, M. globosa was most commonly isolated in total (48 of 180, 26.7%) (Fig. 3). However, among the three body sites, a statistically significant difference was detected only on the forehead (Table 4).
DISCUSSION
Malassezia yeasts are lipophilic fungi and they are considered to be normal flora of the skin, and they are isolated in 75~80% of healthy subjects1,2. Previously named as Pityrosporum in 1889 by Baillon9, Malassezia yeasts are now classified as dimorphic fungi after it was revealed that they harbor hyphae. Even though morphologic variation had been described from earlier times, only M. pachydermatis had been classified as Malassezia species, other than M. furfur. However, as the morphologic, immunologic, physiologic and molecular biological studies have progressed, the need for a new classification system of the species was put forth, and in 1996 Guého et al.10 re-classified the yeasts into seven species (M. furfur, M. obtusa, M. globosa, M. slooffiae, M. sympodialis, M. pachydermatis and M. restricta) based on the morphologic, microscopic, physiologic and molecular biological characteristics. Also, four new species called M. dermatis, M. japonica, M. nana and M. yamatoensis, have recently been introduced from Japan, and in Europe, two additional species called M. caprae and M. equina have been identified, and the yeasts are now classified into thirteen species11-15.
Malassezia yeasts affect the follicles and the keratin layer, where there is an abundant amount of essential free fatty acids and triglycerides16. Malassezia-associated dermatologic disorders can be divided into the following two groups17: the first group includes disorders of skin that are directly caused by Malassezia, and this includes Malassezia folliculitis and pityriasis versicolor. The second category can be described as aggravation of pre-existing skin diseases by growth of Malassezia, and the examples include seborrheic dermatitis, atopic dermatitis and psoriasis.
Malassezia yeasts may turn pathogenic under specific conditions. These conditions include high temperature, high humidity and internal factors such as the long-term use of corticosteroids and immunosuppressants, chemotherapeutic agents, bone marrow transplantation, AIDS, leukemia and diabetes, which elicit the overgrowth of otherwise harmless Malassezia yeasts1,2,18. According to the previous studies, M. globosa is commonly isolated in pityriasis versicolor19-22, and although the reports on seborrheic dermatitis have varied22-24, M. restricta seems to be the most commonly found species in Korea25. The studies on Malassezia folliculitis by Jang et al.26 revealed that M. restricta and M. globosa are the commonly isolated species. Our study revealed that depending on the site, M. restricta or M. globosa is the most common strain that resides in a lesion.
Since Weary27 first reported on the correlation between Pityrosporum and folliculitis and acne, Potter et al.28 presented a detailed report on Malassezia and Malassezia folliculitis. Malassezia folliculitis is a mycotic skin disorder in which an itchy follicular rash and pustules develop on the upper trunk and shoulders where there is abundant secretion by sebaceous glands. It is prevalent during summertime in tropical climates, and a closed environment is a crucial factor for its pathogenesis. Various factors such as the use of tetracyclines or related antibiotics, systemic or local administration of corticosteroids, diabetes, bone marrow transplantation and Cushing syndrome are thought to cause overgrowth of Malassezia yeasts among the resident flora, and this manifests with the aforementioned symptoms1,2. Because rash and pustules may develop on the face and alongside the trunk and shoulders, folliculitis must be differentiated from deep-seated acne. It can be distinguished from acne by the fact that it develops mostly on the trunk and that there are no comedones29.
The diagnosis of Malassezia folliculitis is made by its characteristic clinical manifestations, direct smear microscopy, histopathological examination and the fact that it responds to antifungal therapy. The majority of cases can be treated by local application of antifungal agents, yet some cases may require the oral administration of ketoconazole, itraconazole or fluconazole6.
On comparing the healthy controls and the Malassezia folliculitis patients, five species of Malassezia, namely M. restricta, M. sympodialis, M. furfur, M. globosa and M. dermatis were identified in each group. Also, the most commonly isolated species in the Malassezia folliculitis patients group was M. restricta, while M. globosa was the most common in the healthy control group. The detection rate was the highest in the teens and thirties subgroups in the Malassezia folliculitis group, and in the control group, the thirties age subgroup showed the highest rate.
Among the lesions sampled on the chest, M. globosa was isolated in 13 cases (21.7%), followed by M. sympodialis in 11 cases (18.3%), and on the forehead and cheek, M. restricta was the most predominant species with 17 cases (28.3%) and 12 cases (20.0%), respectively, followed by M. sympodialis with 12 cases (20%) and 10 cases (16.7%), respectively. The outcome differed somewhat from that of the report by Jang et al.26, which stated that M. restricta was found in 4 cases (20%), M. globosa was found in 2 cases (10%) and M. furfur was found in 1 case (5%) on the face, while M. globosa was only predominant with a total of 12 cases (65%) on the neck, chest, flank and sacral area. These disparities may be attributable to the difference in the methods of the two studies, i.e., morphological analysis, such as the size, surface contour, color and shape of the Malassezia colonies, and molecular analysis, such as PCR and RFLP. In addition, other factor such as the differences in the number of subjects, the length of the study, the temperature and the humidity at the time of measurement and various technical factors may also have played roles.
We were able to obtain statistically significant differences between the patient and control groups on the forehead and for the teens and twenties subgroups.
For the results, we identified that the detection rate of M. globosa was lower and that of M. sympodialis was higher, except in the twenties group, as compared to that of the control group.
Although M. restricta was most commonly detected in the Malassezia folliculitis patients, the lower detection rate of M. globosa and the higher detection rate of M. sympodialis suggest that M. sympodialis might possess a pathogenic potential; however, the selection of the subjects and the research methods, along with environmental factors, have to be taken into consideration. According to the report by Akaza et al.30, in which the identification rate of the Malassezia species on the back, upper chest, and neck was compared between 32 Malassezia folliculitis patients (average age: 31±11 years) and 40 controls (average age: 33±7 years), M. sympodialis was detected at a not insignificant frequency in all the specimens. The microflora of the skin, such as M. sympodialis, are known to stimulate keratinocytes to secrete certain specific cytokines and to induce inflammation of hair follicles, thus leading to folliculitis31. Further, Kim et al.32 have reported that M. sympodialis was identified as the main causative agent on the face of a neonate with Malassezia folliculitis. M. sympodialis was found on mycological examination of the folliculitis lesions of renal cell carcinoma patients who underwent treatment with erlotinib, an anticancer agent33, and this further increases the possibility of this strain being pathogenic. However, considering that there have been reports, such as those by Crespo and Delgado6 and Rhie et al.34, that the main causative agents of Malassezia folliculitis are normal flora of the skin other than M. sympodialis, and that other strains such as M. furfur, had been isolated from the neonatal Malassezia folliculitis lesions on the face3, and given the outcome of this study in which the rate of identifying M. sympodialis in the twenties control group was lower than that in the patient group, the possibility of this strain being pathogenic may be questioned.
Therefore, in order to identify the Malassezia species that play a major role in the pathogenesis and aggravation of Malassezia folliculitis, a large-scale quantitative analysis in future studies conducted on a larger patient pool and more variable sites of lesions may be needed.






 XML Download
XML Download