

 PDF
PDF ePub
ePub Citation
Citation Print
Print


서론
종양 세포 중 높은 악성도를 가진 일부의 암세포에 의하여 종양의 성장이 유지된다는 개념에서 암줄기세포가 제안되었고, 이러한 세포들은 많은 백혈병과 고형 종양에서 분리되어 왔다. 암줄기세포는 암세포 계층 구조의 최상위를 구성하며 최근의 연구에서는 이러한 계층 구조가 유전적으로 다양하며 종양 내부에서 서로 연관성은 있으나 뚜렷이 구분는 다양한 하위 클론이 존재하며 종양 내부에서 서로 경쟁하고 있다는 것이 발견되었다. 암줄기세포의 개념은 종양의 재발과 전이의 매우 복잡하고 역동적인 과정을 더욱 잘 반영할 수 있는 모델로 진화하여 왔다.
암줄기세포의 개념
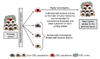
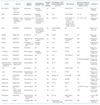
종양조직 내 일부 세포들만이 종양전파 능력이 있다는 암줄기세포의 존재에 대한 가설은 놀랍게도 수십 년 전부터 보고되어 왔었다. 1937년 Furth 등[1]에 의해 한 개의 세포에 의해 생쥐모델에서 백혈병 전파가 가능함이 보고되었고, 1960년대에 Bruce와 van der Gaag [2]가 종양조직 중 일부 세포만이 종양을 형성할 수 있음을 이미 보고하였다. 그러다가 1997년 Bonnet과 Dick [3]에 의해 급성골수구성백혈병에서 그 존재가 명확히 입증되어 많은 사람들의 관심을 끌기 시작했고, 이후 뇌종양(2004년), 대장암(2007년), 암세포주들에서도 그 존재가 보고되었다[4] (Table 1). 인체의 각 조직에 줄기세포가 존재하여 장기를 유지시켜 주듯이, 암조직 내에도 1-2% 정도의 소수 암줄기세포가 있어서, 정상줄기세포처럼 자기재생능력도 있고, 다른 세포로 분화할 수 있는 능력을 지니고 있다는 것이 암줄기세포의 기본 개념이다[25]. 또 정상조직에서 감염이나 손상 등에 의해 세포손실이 생기면 줄기세포가 분열하여 재생 촉진을 유지하는 역할을 하듯이, 암조직 세포들도 계층적인 조직 구성을 지니고 최상부에 암줄기세포가 자리잡고 있다고 생각되고 있다[26] (Figure 1). 현재까지 생식세포암[27], 백혈병[3], 유방암[9], 뇌암[11], 대장암[12], 췌장암[24], 흑색종[19] 등의 암에서 암줄기세포에 의해 지배되는 계층구조가 보고되었다. 암줄기세포의 유래에 대해서는 줄기세포에서 기원했을 수도 있고, 일부 분화된 암세포가 줄기세포의 형질을 획득하여 생길 수도 있다고 생각되고 있는데, 현재로서는 아직 명확한 결론은 나지 않은 상태이다.
암줄기세포의 존재는 종양조직에서 종양세포를 단일세포로 분리하여 각각의 세포를 따로 따로 동물에 주입하여 종양형성능력을 관찰하여 입증되었다. 종양조직에서 종양세포를 분리하여 암줄기세포 표지자로 염색한 후 유세포분석기를 이용하여 암줄기세포를 단일세포 상태로 분리하여 동물에 주입하고 종양형성을 관찰하였다. 종전에는 종양조직 내 세포를 분리하여 충분량의 세포 수를 동물에 주입시켜 종양형성을 관찰하였으므로 암줄기세포와 나머지 암세포들이 혼재되어 암줄기세포에 대한 판단이 사실상 불가능하였었다. 암줄기세포의 생태를 연구하기 위해서는 단세포조작(single cell manupulation)이 필수적이며, 이러한 면에서 앞으로의 암줄기세포에 대한 연구는 과학적인 면보다는 실험기술, 즉 기술(art)에 달려 있다고 이야기되기도 한다.
암줄기세포의 특성
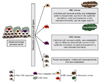
암줄기세포의 조건으로는 첫째, 종양을 형성할 수 있어야 하며, 둘째, 생체 내 종양전파(in vivo propagation)가 가능하여야 하며, 셋째, 다양한 형질을 지닌 세포들로 분화가 가능한 세포들을 만들어 낼 수 있어야 한다. 분화된 자손세포와 암줄기세포가 다른 점은, 암줄기세포는 무제한적인 자가 갱신 분열을 할 수 있다는 것이다. 인체 내 정상적으로 존재하는 정상줄기세포와 암줄기세포와 비교하여 보면, 비슷한 점은 4가지로 요약할 수 있다. 첫째, 두 가지 세포 모두 비대칭분열(asymmetric division)을 할 수 있다. 대칭분열(symmetric division)은 자신과 동일한 세포를 생산해내는 분열을 의미하며 비대칭분열이란 자신과 동일한 세포와 일부 분화한 세포를 동시에 생산해내는 것을 일컫는다(Figure 2). 둘째, 정상줄기세포는 재생을 통해 개체의 장기를 일생 동안 유지하는 역할을 하는데 암줄기세포도 역시 종양을 증식시키기 위한 자기재생능력이 있다. 셋째, 분화능력이 있다는 점이다. 정상줄기세포는 장기에 필요한 세포로 분화하며, 암줄기세포 역시 분화하여 다양한 표현형을 보이는 장기의 종양세포를 생산해낸다. 넷째, 정상줄기세포나 암줄기세포 모두 유사한 세포 내 신호전달 경로에 의해 재생이 조절된다.
그렇다면 정상줄기세포와 암줄기세포가 다른 점은 무엇인가? 정상줄기세포에서 자기재생에 관여하는 신호전달경로가 정상적으로 조절될 경우 정상조직이 유지되며, 신호전달경로에 이상(dysregulation)이 생길 경우 암줄기세포로 된다고 생각되고 있다. 우리 몸에서는 하루에 1,011개의 세포가 새로이 형성되는데 이론적으로 106개 세포 중 하나는 돌연변이가 발생하게 된다. 그렇다면 하루에도 수백만 개의 돌연변이를 지닌 세포가 생성되는데 이들 세포가 암세포가 되지 않는 이유는 암발생은 다단계과정(multistep process)에 의해 발생하기 때문에 단일돌연변이만으로는 암발생에 불충분하기 때문이다. 또 우리 몸의 대부분의 세포들은 분열이 불가능한, 즉 이미 완전히 분화된 세포들로 대부분 이루어져 있으며, 돌연변이를 지닌 세포가 있더라도 대부분의 세포가 여러 단계의 돌연변이 변화를 거칠 만큼 오래 살지 못한다는 점이다.
암줄기세포의 개념 변화와 새로운 동적 개념
과거에는 종양세포 중 높은 악성도를 가진 소수의 세포에 의하여 종양의 성장이 유지되고, 그 세포들이 바로 암줄기세포라고 생각했었지만, 지난 10년간 연구에 의하면, 암줄기세포는 종양조직 세포중 최상위 계층에 위치하고, 유전적으로 다양한 암세포들이 그 하위 구조를 이루고 있다고 생각된다. 이들 다양한 세포들은 연관성은 있지만 서로 뚜렷이 구분되며, 종양 내부에서 치열한 경쟁하고 있다. 즉, 종양은 동일 특성을 가진 세포집단이 아니라 매우 다양한 세포로 구성되어 있고, 암줄기세포의 개념이 종양의 재발과 전이라는 매우 복잡하고 역동적인 과정을 더욱 잘 설명해 줄 수 있는 모델이라 여겨지고 있다. 암조직을 이루는 세포들의 성격을 세 가지로 정리하면 다음과 같다. 첫째, 종양은 유전적으로 불안정하여, 초기 단계, 재발 시, 혹은 진행 병기에서 서로 아주 다른 유전적 차이를 보일 수 있다. 초기에는 아주 적은 숫자의 암줄기세포들이 종양을 유발시키지만, 진행하는 동안, 돌연변이에 의한 새로운 암줄기세포 클론이 생겨날 수 있다. 항암치료 혹은 표적치료를 통하여 대부분의 종양세포가 제거된 후에는, 소수의 암줄기세포 클론만이 생존하여 증식하고, 재발한 종양은 주 구성성분이나 성격이 크게 달라질 수 있다(Figure 2A). 둘째, 서로 다른 유전적으로 특이한 암줄기세포 클론들은 서로 경쟁하여, 분열능력이 강한 클론이 선택되고 분화능은 소실되게 된다. 결국 진행된 종양에서는 계층적 구조가 평면화되고, 과거 확률적 모델에서 제시된 것과 유사한 상황이 되게 된다(Figure 2B). 셋째, 암줄기세포의 바이오마커는 한가지로 고정되어 있는 것이 아니라 암의 병기마다 다양하게 변할 수 있다(Figure 2C). 그 이유는 앞선 설명처럼 암이 진행하면서 그 구성 세포들의 성격이 달라지기 때문이다. 따라서 기존 보고된 암줄기세포 바이오마커들은 특정 종양 병기에서만 유효할 수 있고, 따라서 각각의 경우에 따라 기능성 분석과 함께 진행병기마다 다시 검증되어야 의미가 있다.
이러한 사실의 임상적 의미는 이러한 동적 암줄기세포 모델이 진행된 병기의 종양이나 전이를 표적으로 하여 치료하는데 유효할 수 있다는 것이다. 한 종양 내 모든 클론에서 공통적으로 존재하는 돌연변이를 게놈 염기서열분석 등의 방법을 이용하여 확인해야 한다. 공통적인 돌연변이에 의해 조절 작용에 이상이 생긴 관련된 신호전달 기전을 표적으로 효율적인 치료를 시도할 수 있다. 이러한 전략으로 부분적 종양 클론만 제거하지 않고, 진행된 병기의 암이나 전이부위에서 나타나는 다양한 암줄기세포를 제거하는 표적치료를 할 수 있다.
한 종양 내에서 유전적으로 서로 다른 여러 종류의 암줄기세포 클론이 공존한다
췌장암과 신장암에서 원발종양과 전이종양간의 염기서열 분석을 하여 보았더니 예상과 달리 이질성들이 발견되었다[2829]. 이러한 이질성들은 암줄기세포가 유전적으로 불안정하다는 사실에서 초래된, 어찌 보면 당연한 현상이라 할 수 있다. 실제 대장암에서 그러한 유전적 불안정성이 보고되었고 부모 암줄기세포 클론으로부터 새로운 암줄기세포가 형성되는 현상이 보고되었다[30]. 또, 급성림프구성백혈병에서도 질환의 진행 동안과 이종이식 쥐에서의 이식 후에 백혈병 줄기세포의 유전적 구성을 조사하였더니, 여러 종류의 백혈병 줄기세포 클론들이 한 환자 내에 공존하고 있는 것이 보고되었다. 암이 진행하는 동안, 혹은 항암치료 후에 그때마다 다른 암줄기세포 클론이 힘을 얻게 되고, 원발종양과는 다른 성격의 종양을 형성할 수 있다[31]. 약물 감수성을 보이는 암줄기세포들이 표적치료로 감소한 후에는, 이미 존재하고 있던 저항성을 가진 소수클론이 우세해지면서, 마치 치료에 대해 내성을 획득한 것처럼 보일 수도 있다. 어쩌면 종양은 꼭지점이 하나인 계층구조에 의해서 나타내어 질 수 없고, 여러 개의 꼭지점을 가진 구조에 더 가까울 수 있다. 암줄기세포 클론의 꼭지점이 표적치료에 의하여 잘려 나간다 하여도, 다른 꼭지점이 우세하여져서 종양을 다시 형성할 수 있다는 것이다. 따라서 시시각각으로 변하는 동적 계층구조가 하나의 암줄기세포 클론과 그 자손세포 내(암줄기세포 클론 계층구조 내)에 존재할 수 있지만, 서로 경쟁하는 유전적으로 다양한 암줄기세포 클론들(암줄기세포 클론 계층구조 간) 간에도 존재할 수 있다(Figure 2A).
암줄기세포는 반드시 드물게 존재할 필요는 없다
암줄기세포는 반드시 드물게 존재해야 할 필요는 없다. 사실, 암줄기세포가 풍부한 종양은 0.2%에서 82.5%까지 매우 다양한 비율을 보이는 것으로 보고되었다(Table 1). 또한, 종양의 진행병기마다 달라서 진행에 따라 빈도가 증가할 수 도 있다. 저등급과 고등급의 유방암 종양의 제한 희석 검사 지난 몇 년 동안 암줄기세포는 드물어야 한다는 생각이 암줄기세포 가설에 대한 의문들을 일으켜왔다. 예를 들어 흑색종 3기 혹은 4기에서 종양을 생성할 수 있는 세포는 매우 많아서, 종양의 30%까지 이르기도 한다[21]. 어떤 사람들은 이런 현상을 두고 흑색종은 암줄기세포 모델을 따르지 않고, 종양형성능이 암조직 세포 중에 무작위로 흩어져 있다는 확률적 모델을 따른다고 해석하기도 하였다[32]. 그러나 다른 설명 또한 가능하다. 진행된 병기의 흑색종은 서로 다른 표면 표지자를 발현하는 여러 개의 구분되는 매우 악성도가 높은 암줄기세포 클론으로 구성되어 있는데, 분화능이 거의 남아 있지 않은 암줄기세포 클론이 선택되고 농축되어 있는 상태라는 설명도 가능하다. 이런 말기 암 상태에서는 처음의 종양 계층구조가 점차 사라지고 이질적인 암줄기세포 클론이 독점적으로 자리를 차지한 상황이 만들어지게 된다(Figure 2B).
암줄기세포는 어디에서 유래하는가?
암줄기세포의 유래에 관하여 분열능력이 높은 세포가 돌연변이에 의해 성상이 달라지면 암줄기세포로 된다는 학설이 "stem cell theory of cancer"이다. 조혈모세포의 예를 들어 보자. Long term hematopoietic stem cell은 symmetric division과 모든 계열의 혈액세포로 분화 할 수 있는 assymetric division을 할 수 있는 능력을 지니며, short term hematopoietic progenitor cell을 거쳐 특정세포계열로만 분화가 가능한 수임조혈모세포(committed multipotent progenitor cell)로 분화하면 이를 거쳐 특정혈액세포로 성숙하게 된다. 이러한 조혈계층 형성의 초기 단계들 중, 조혈모세포 수준 혹은 좀 더 진행된 수임조혈모세포 수준에서 돌연변이나 유전자 이상이 생기면 암줄기세포로 이행하는 것으로 생각되고 있으며 이러한 개념은 유방암 줄기세포에서 마찬가지로 적용되고 있다. 여기에서 유의하여야 할 점은, 만성백혈병이 발생하는데, 이들 BCR-ABL 융합유전자는 조혈모세포 유사 만성골수성백혈병 세포에 존재하고 있음으로 보아, 질환의 기원이 되는 세포가 정상 조혈모세포이라고 생각되고 있다. 그러나 모세포 위기의 진행된 단계의 백혈 암줄기세포 개념은 세포의 기원과는 따로 분리해서 생각하여야 한다는 것이다. 암줄기세포는 기능적인 특성으로 정의되지만, 종양세포의 기원은 암발생 돌연변이가 처음으로 발생한 세포의 유형을 의미한다. 그러나, 이렇게 기원한 세포가 반드시 바로 암줄기세포의 표현형을 획득하지는 않는다. 암줄기세포는 줄기세포 단계가 아닌 분화된 전구세포 단계에서 줄기세포 형질을 획득하여 생길 수도 있다. 즉, 초기의 유전 돌연변이를 획득한 줄기세포에서 유래된 전구세포들이 분화하면서 더 많은 돌연변이를 획득하여 새로운 암발생 혹은 재발을 일으킬 수 있다. 첫 돌연변이에 대한 기원 세포는 줄기세포이지만, 암형성 클론을 형성하는 암줄기세포는 더 분화된 전구세포임을 의미한다. 이러한 시나리오는 만성골수성백혈병에서 보고되었다. BCR-ABL 융합단백이 만들어지면 병 줄기세포는 과립구계/대식구계 전구세포와 유사한 상태에 있다고 보고되었다[33]. 급성골수성백혈병에서도 AML-ETO 융합단백이 조혈모세포(이 질환의 기원이 되는 세포)에서 나타나지만, 실험실적인 콜로니 형성 검사에서 기능적인 백혈구 줄기세포가 Thy1- 전구세포 단계에 있음이 보고되었다[34]. 급성전골수구성백혈병(acute promyelocytic leukemia)에서는 MLL-AF9 융합단백이 조혈모세포에서는 발견되지 않았지만[35], 쥐의 과립구계/대식구계 전구세포에 도입되었을 때 백혈병을 유발할 수 있었으며, 이 사실은 기원 세포와 백혈병 암줄기세포가 모두 조혈모세포가 아닌 전구세포라는 것을 의미하고 있다[36].
암줄기세포를 찾아낼 수 있는 바이오마커는 무엇인가?
암줄기세포의 개념은 치료효과를 평가하는 중요 인자 중 하나로 종양세포 수의 감소보다는 치료 후 잔류 암줄기세포가 얼마나 남아있나 하는 것이다. 따라서, 각 종양 유형에 대한 신뢰성 있고 특이적인 암줄기세포 바이오마커를 찾는 것이 관건이라 할 수 있겠다. Bonnet과 Dick [3]은 급성골수성백혈병의 백혈병 줄기세포가 정상 조혈모세포와 유사하게 CD34+/CD38- 세포라 보고하였지만, 개인마다 많은 이질성이 있다고 알려져 있다[7]. 고형암에 대해서는, 2003년 Al-Hajj 등[9]이 폐삼출액과 종양 검체에서 ESA+/CD44+/CD24low-neg 세포군이 유방암 암줄기세포라 보고하였고, Table 1에서와 같이 많은 바이오마커들이 보고되었다. 보고가 많아짐에 따라, 같은 종양 유형 내에서도 때로는 서로 다른 표지자를 보이는 암줄기세포가 존재한다는 것이 알려졌다. 이러한 불일치는 방법상의 차이가 원인이 될 수도 있고[37], 여러 종류의 암줄기세포 클론들이 일차적인 종양 내부에서 공존하고 있기 때문에 연구그룹들이 서로 다른 암줄기세포군을 검출하였을 수도 있다[38]. 실제로, 암의 실체 자체는 거의 균일하지 않고 이질적이다. 예를 들어 지난 몇 년간, 바이오마커와 임상적 접근을 동반하여 다양한 게놈 차원의 유전자 발현 프로파일 분석으로 유방암은 점차 세분화된 분자적 소분류로 나뉘게 되었다[39].
이들 암줄기세포 바이오마커들은 잔존 암줄기세포를 찾는 목적 이외에도 표적치료에도 이용될 수 있다. CD44는 막관통당단백으로 세포부착에 관련하며 많은 암줄기세포에서 발현되는데(Table 1), 이에 대한 CD44 항체가 암줄기세포와 미세환경과의 상호작용을 억제하여 급성골수성백혈병 줄기세포의 증식을 억제한다는 보고가 있었다[40]. 그러나, 정상 줄기세포에서도 발현되는 CD44에 대한 치료는 심각한 독성을 유발시킬 수 있다. CD123 (interleukin-3 수용체 α 사슬)도 이종이식 쥐에서 차단항체를 이용하여 급성골수성백혈병 암줄기세포를 성공적으로 제거할 수 있었으나[41], 정상 조혈모세포에서도 발현되기 때문에 심각한 부작용의 위험을 안고 있다. 암줄기세포에서 발현되는 ABC 수송체에 대한 표적치료 또한 암줄기세포 억제효과는 있으나, 이 수송체가 혈액-뇌 장벽의 유지 및 성인 줄기세포의 유지에 중요한 역할을 한다는 것이 밝혀지면서 연구가 보류되었다[42].
암줄기세포의 존재는 암치료에는 어떠한 의미를 지니는가?
암줄기세포는 다른 암세포와는 달리 휴지기 상태에 있으며 따라서 형태도 비교적 얌전하다. 현재의 항암 치료방법은 모든 암세포가 분열능력 및 전이능력이 있다고 가정하고 치료를 하는 것이다. 치료를 하면 종양조직은 작아지고 마치 항암제에 잘 반응하는 듯 보이지만 소수 잔존하는 암줄기세포에 의해 다시 재발하게 된다. 암줄기세포는 다른 세포와는 달리 세포주기 중 휴지기에 있으므로 항암제의 공격에 저항하여 소수로 살아남아 있다가 양호한 환경이 되면 다시 빠른 속도로 증식하여 임상적으로 관해가 온 것처럼 보이는 환자에서 재발을 일으키게 되는 것이다[43]. 예를 들어 유방암의 경우 외과적 절제 및 성공적인 항암치료 후 10년간 재발 없이 지내다가도 아주 작은 임파절 전이가 발견되면서 아주 빠른 속도로 전신전이가 진행되는 예를 볼 수 있다. 이는 오랫동안 휴지상태로 존재하던 암줄기세포가 시간이 흐르면서 활성화 시키는 발암자극(oncogenic hit)을 받게 되면 동면상태에서 벗어나 분열할 태세를 갖추게 되고 여기에 다시 두 번째, 세 번째의 다단계 발암자극을 받게 되면 세포주기 내로 들어가 아주 빠른 속도로 분열하게 되기 때문이다.
휴면 또는 느린 세포주기 순환 상태의 암줄기세포
일부 암줄기세포에서 성인 줄기세포와 유사하게 동면 또는 느린 세포주기 상태와 활성 상태를 순환한다는 것이 보고되었다[44]. 암줄기세포가 휴지기 상태에 있으면 세포주기를 표적으로 하는 항암제를 피해 나갈 수 있으며, ABC transporter 등의 펌프가 잘 발달한 세포이다. 따라서 독성이 있는 물질이 암줄기세포 내로 침투하여 축적되지 못하도록 보호되며, 암줄기세포의 니쉬(niche)에 의해 보호되어 돌연변이로부터 보호된다는 특성이 있다. 이러한 특성들은 개체가 살아나가는 동안 노출되는 여러 독성물질이나 열악한 환경으로부터 줄기세포를 보호하기 위한 지극히 타당한 특성이지만, 암줄기세포 역시 이러한 특성을 가지고 있으므로 항암제를 투여하거나 방사선을 조사하더라도 암줄기세포 이외의 나머지 세포들은 잘 죽지만 암줄기세포는 살아남게 된다. 놀라운 사실은 암줄기세포는 평상시에는 휴지기 상태이지만 특정 상황에서 일단 활성화되면 빠른 속도로 세포주기로 들어가 증식할 수 있는 능력을 지니고 있다는 점이다. 이렇게 휴면 중인 암줄기세포의 재활성화가 치료 완료 후에 수 십 년 후의 종양 재발을 유도할 수 있다는 가설도 제시되었다[45].
항암치료에 대한 저항성을 보이는 암줄기세포
생리적인 줄기세포와 유사하게 암줄기세포는 항암치료에 대하여 높은 저항성을 보인다. 예를 들어, 유방암 항암치료 후 종양세포의 상당 부분이 제거된 종양조직에 암줄기세포가 축적되는 것이 발견되었다[46]. 비슷하게 만성골수구성백혈병에서도 imatinib과 같은 tyrosine kinase 억제제 사용으로 완전 분자 관해에 도달한 후에도 약물저항성 줄기세포가 남아 치료가 중단된 동안 재발을 유발하였다[47]. 약제 내성 암줄기세포에 대한 다른 증거는 5q 소실을 동반한 골수이형성증후군에서 보고되었다. 대부분의 종양세포들은 면역조절약물인 lenalidomide에 의하여 효과적으로 치료되어 임상관해 및 세포유전학적 관해 상태에 도달하지만 대부분의 환자들은 남아 있는 약물저항성 암줄기세포의 성장에 의하여 재발하게 된다[48]. 현재 암줄기세포의 약물저항성의 원인은 세 가지로 요약되고 있다. 첫째, 암줄기세포는 다약재내성 수송체의 발현을 통하여 약재를 유출시키는 능력이 있다[49]. 둘째, 알데하이드의 산화촉매와 관계된 세포 내 효소인 aldehyde dehydrogenase 1 (ALDH-1)이 여러 종류의 암줄기세포에서 특이적으로 활성화되어 있다[10]. 실제 ALDH-1의 활성도는 유방암 전이세포에서 현저히 증가되어 있었고, cyclophosphamide에 대한 내성을 초래한다고 보고되었다[50]. 셋째, 암줄기세포는 증가된 DNA 복구능력을 지니므로 전리방사선치료 등의 유전독성치료에 대해 세포독성 효과를 상쇄할 수 있다고 생각된다. 이는 CD133 발현 및 비발현 교모세포종(glioblastoma)에서 증명되었다[51]. 넷째, 암줄기세포는 자유라디칼 제거 작용이 항진되어 있어 방사선에 의하여 생성된 활성산소기를 상쇄할 수 있으며, 이는 쥐와 사람의 유방암줄기세포에서 보고되었다[52]. 이러한 능력은 암줄기세포를 활성산소에서 유도된 DNA 손상으로부터 선별적으로 보호하며, 암줄기세포의 방사선치료에 대한 저항성을 잘 설명하고 있다. 다행히도, 약물 감수성 종양 세포주에 대한 생체 외 연구에서는 암세포는 일시적이며 가역적으로 약물 저항성을 획득할 수 있어, 약제 내성이 항상 안정적인 특성은 아니라는 희망적인 연구보고도 있다[53].
암줄기세포는 어디에 위치하는가? 줄기세포의 미세환경과 니쉬
줄기세포의 니쉬(stem cell niche)는 줄기세포의 기능을 유지하고 보호하기 위한 환경으로 줄기세포와 부착물질(adhesion molecule)을 통해 물리적으로 부착이 가능한 위치에 존재한다(Figure 3). Stem cell hh, Wnts, bone morphogenetic proteins, fibroblast growth factors, Notch 등의 외부 시그날을 통해 줄기세포의 사멸, 분열, 분화를 조절한다. 평상시에는 줄기세포를 slow cycling, self-renewing, undifferentiated state로 유지하고 있으나, 암세포에서는 세포를 분열시키려는 힘이 더 우세해져서 세포주기 내로 들어가 분열하게 된다. 알려져 있는 줄기세포의 니쉬로는 장상피세포(intestinal epithelium stem cell niche), 모낭의 hair follicle epidermal stem cell niche, 골수의 조혈모세포 등이 있다.
줄기세포 미세환경
종양의 미세환경은 다양한 면역세포와 기질세포, 그리고 세포 외 구성요소로 이루어져 있다. 암줄기세포의 기능적 특성은 바로 이러한 "니쉬"로 명명된 미세환경에 의하여 유지될 수 있다. 예를 들어, 혈관내피세포는 쥐 이종이식 모델에서 교모세포종 암줄기세포의 자가 갱신능을 유지하고, 암 형성 능력을 촉진하고[54], 저산소증은 hypoxia-inducible factor 2-α를 통하여 교모세포종 암줄기세포의 자가갱신능과 암 형성 능력을 증가시킨다[55]. 더 나아가, interleukin 6와 같은 종양에 침윤한 면역세포에 의하여 분비되는 염증관련 분자들은 장염 관련 암줄기세포의 증식을 촉진한다고 보고되었다[56]. 암줄기세포 미세환경은 암줄기세포의 특성을 조절할 뿐 아니라 암줄기세포가 아닌 세포에 암줄기세포의 특성을 부여하기도 한다. 즉, 암줄기세포를 조종하는 능력도 있으면서, 암줄기세포를 만들 수도 있다고 할 수 있다. 예를 들어, 종양 관련 근섬유모세포는 간세포 성장 촉진인자의 분비를 통하여 대장 암줄기세포의 갱신능을 증진시키고, 한편으로 분비물질을 통해 암줄기세포가 아닌 세포들의 생체 내 암 생성능력을 강하게 활성화 시키기도 한다[57]. 정상 줄기세포의 미세환경은 휴면상태의 유지와 스트레스를 유발하는 자극에 대한 저항성에 중요한 역할을 하는 것으로 알려져 있으나, 그러나 종양 미세환경이 암줄기세포의 기능에 미치는 세포 수준과 분자 수준의 영향에 대해서는 아직도 연구가 더 필요하다.
암줄기세포를 표적으로 하는 새로운 전략
최근 연구들은 표적세포에 대해 양방향 특이 항체 혹은 삼방향 특이 항체를 제작하여 표적치료를 시도하고 있다. 예를 들어, 항 CD3항체를 통하여 T 세포를 인식하면서, 동시에 EPCAM이나 HER2와 같은 다른 항원을 통해 암세포도 인식하는 방식이다. 이러한 방법으로 원하는 세포의 특성을 잘 선별하여 적중시키게 되면 정상 세포를 보존하면서 원하는 세포만 표적살상 할 수 있게 된다[58]. 또, 초고속 약물스크리닝 기법으로 암줄기세포를 선택적으로 억제하는 약물, salinomycin과 같은 화합물을 발견하게 되었다. 이러한 항생제 약물은 잠재적인 유방암 암줄기세포를 표적으로 하며, 경로 억제를 통하여 만성림프구성백혈병의 암줄기세포를 억제할 수 있다고 한다[59].
마지막으로, 암줄기세포의 특성이 잘 연구되면, 곧 새로운 표적 전략의 발전으로 이어질 수 있다. 예를 들어, 교모세포종 줄기세포는 산화질소 합성효소-2(nitric oxide synthase 2, NOS2)를 통하여 산화 질소를 생산한다는 측면에서 종양 형성능이 없는 대응되는 정상세포와 기질적으로 구분이 되는데, 이종이식 교모세포종 모델에서 이러한 NOS를 억제하며 종양을 억제시키는데 성공하였다[60]. 비슷하게, 글라이신 탈카르복시화 효소 활동은 비소세포 폐암 암줄기세포에서 중요한 것으로 밝혀져, 글라이신 대사과정이 새로운 항 암줄기세포 표적이 될 수 있을 것으로 생각된다[18]. 그러나 이 두 경우 모두, 일반적인 치료 독성은 아직 충분히 평가되지 않았다. 암줄기세포와 정상줄기세포의 분자 발현 유사성에 의하여 발생하는 치료의 독성이 이러한 치료의 임상적 사용을 방해하는 주요 장애물이라 생각된다.
암줄기세포와 전이
상피-중간엽 전환(epithelial-mesenchymal transition, EMT)은 정상적 배아발달 과정에서 발생하는 현상으로, 세포가 상피(epithelial)성 세포 표현형을 상실하고 이동성이 높은 중간엽(mesenchymal)성 세포 표현형으로 전환하는 과정으로 암전이에도 관여하는 것으로 알려져 있다. 아직까지, 암의 전이 개시세포(metastasis-initiating cell, MIC)가 확실하게 확인이 되지는 못했지만, 전이 개시세포가 암줄기세포의 하위 집단 내에서 존재한다는 보고가 있다. 암줄기세포는 멀리 떨어져 있는 장기에 이차적인 종양을 만들기 위해 종양 개시 능력을 필수적으로 가지고 있으며, 이들은 EMT 표지자를 발현하고 있다[61]. EMT 표지자를 발현하고 있다는 사실은 그 세포가 이동할 수 있고 전이 개시 활동의 가능성을 가진다는 것을 의미한다. 전이 개시세포(MIC)는 암 초기에 파종될 수도 있고, 말기에 파종되는 암줄기세포 클론에서 생성될 수도 있다. 실제 차세대 염기서열 분석기법으로 원발췌장암과 전이암을 비교하여 보니, 모두 이질적인 클론으로 구성되어 있지만, 원발 종양과 비교하여 추가적인 개시 돌연변이들이 전이-개시 클론에 존재한다는 사실이 보고되었다[62]. 전이는 멀리 떨어져 있는 조직에 원발종양세포를 성공적으로 전파하는 것이므로, 전이 개시세포(MIC)는 전파된 종양세포 중에서 발견된다. 여기에 상피암세포 환자의 혈액이나 골수에서 발견되는 순환종양세포(circulating tumor cell, CTC)나 파종종양세포(disseminated tumor cell, DTC)를 포함된다. 유방암의 경우, 다양한 비율의 CTC와 DTC가 면역화학염색을 통하여 암줄기세포 표현형을 나타내는 것이 보고되었으나[63], 상피암 줄기세포를 검출하는 방법에 대해서는 논란을 안고 있다. 대개의 연구에서 epithelial cell adhesion molecule (EPCAM과 cytokeratine (CK)를 상피암세포 선택 표지자로 사용하는데, 실제 이런 표지자들은 EMT를 통한 파종에서는 그 발현이 감소할 수 있기 때문이다. 이를 밝히는 실험으로 췌장선암 셍쥐모델의 암줄기세포 분석을 들 수 있다. 생쥐모델 췌장암 암줄기세포의 40% 이하에서만 CK19 혹은 EPCAM을 발현하였고, 이는 많은 암줄기세포가 EMT 과정을 거치며 상피 표지자의 발현을 잃어버리게 된다는 것을 의미한다. 파종성 암줄기세포가 정말로 전이 개시에 관여하는지, 만약 관여한다면, 어떤 암줄기세포 하위 집단이 이에 관계하는지 밝히기 위해서는, 이종이식 실험을 이용한 인간 CTC와 DTC의 여러 다른 하위집단들에 대한 기능성 체내 연구가 필요하다. 그러나 현실적으로 이러한 연구들은 적은 숫자의 암세포를 이용한 전파실험 기법이나 전이판독 검사기술의 개발이 뒷받침되어야 하므로 고난이의 기술을 필요하다는 것이 걸림돌이다.
전이 개시세포 모델
전이 개시세포는 말기 단계 암줄기세포 클론의 하위 집단에서 발생하는 것으로 생각되므로, 말기 종양의 암줄기세포과 그 특성과 비슷하다. 이 세포들은 암줄기세포의 기본 특성과 함께, 전이 능력을 가지고 있고, 전신 순환계에서 생존할 수 있어야 하고, 전이된 곳에서 혈관 외로 유출될 수 있어야 하며, 새로운 환경에서 뿌리를 내리고 성장할 수 있어야 한다. 전파되는 동안 여러 종류의 전이성 클론들이 공존할 수 있으며, 각 클론들은 부위별 특이한 이동과 파종 능력을 지니고 있는 것으로 알려져 있다(Figure 3. 실제로, Bos 등[64]은 전신 전이가 있는 유방암 환자의 암조직에서 뼈, 폐, 혹은 뇌로 전이를 할 수 있는 능력이 서로 다른 세포들이 구분되어 존재한다고 보고하였다. 이러한 전이능력의 차이는 아마도 전이개시세포들이 전이 미세환경을 형성하는 능력 때문이라고 추측된다.

 XML Download
XML Download