

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Fusarium fungi is a prevalent toxin-producing mold that produces various mycotoxins including trichothecene mycotoxins (T-2 toxin, deoxynivalenol (DON) and HT-toxin), zearalenone (ZON) and its 2 metabolites (α-zearalenol and β-zearalenol). These mycotoxins are characteristically stable under changing environmental conditions and have been shown to cause a variety of toxic effects in experimental animals, livestocks, and humans.
Deoxynivalenol, one of the most widely spreading contaminants in food and feed, can induce both toxicologic and immunotoxic effects in a variety of cell systems and animal species. For example, DON is cytotoxic to reticulocytes, fibroblasts and lymphocytes [14,18], and the cellular toxicity appears to be mediated by the inhibition of protein synthesis [18]. Deoxynivalenol inhibits cell division, RNA/DNA synthesis and apoptosis [13].
Zearalenone and its metabolites disrupt reproductive processes by mimicking the action of estradiol-17β [6]. In mature and cyclic sows, ZON causes multiple reproductive dysfunctions (25-50 mg/kg ZON added to the diet of pregnant sows causes smaller litters with smaller offspring) [3].
The published critical values of DON and ZON for farm animals are 1 mg DON/kg and 0.25 mg ZON/kg for starting and finishing pig diets and 0.05 mg/kg for pre-pubertal female breeding pigs [2]. However, DON and ZON reportedly caused detrimental effects in farm animals at lower values than the published ones. To date, there is no report of whether DON and ZON cause any detrimental effects on the immune system of pigs. Therefore, the aim of this study was to investigate the effects of DON and ZON combination on the physiological functioning of pigs when the defined concentrations of DON and ZON were incorporated into feed.
Materials and Methods
Animals and treatments
A total of 24 weaning female piglets (~6 kg BW) were obtained from a classical swine fever (CSF) virus-free breeder farm. Four pigs were housed in 1 pen, 3 pens for each group. Animals were allowed to acclimate for 2 weeks to their new housing at 22-24℃ with negative-pressure ventilation before treatments. Pigs were fed experimental diets for 6 weeks.
The 24 piglets were divided into 2 groups, a control group fed a diet free of mycotoxins, and a toxin group fed a diet containing 1 mg/kg DON and 250 µg/kg ZON. The basal diet (Table 1) was primarily based on corn with soybean meal and was formulated according to NRC requirements (1998). During the experiment, feed and water were provided ad libitum through the 6-week experimental period. The ethical guidelines for animal protection rights were observed.
Mycotoxins
DON and ZON, kindly provided by GTI GmbH (IFA Tulln, Austria), were fermented in wheat and barley by a fungal inoculation procedure. The content of mycotoxins incorporated in different treatments was sampled and analyzed by HPLC methods [17]. For the mycotoxin assay in feed, less than 0.01 mg/kg DON and 10 µg/kg ZON in the control group, 1.03 mg/kg in the DON group and 258 µg/kg ZON in the toxin group, were detected.
Immune function evaluations
Pigs were vaccinated s.c. with 1 dose of CSF vaccine at the beginning of the experiment, and received a booster 2 weeks later. Blood samples were collected from the vena cava of all the pigs on d 1, 14, and 28 after first vaccination. The antibody titers for CSF were measured by the anti-CSF antibody ELISA kit (Idexx, USA).
Blood was collected from all the pigs before slaughter on d 42 of the experiment for blood biochemical parameter assays. After centrifugation at 3,000 × g for 10 min, the sera were collected for determination of total protein, albumin, globulin, γ-glutamyltransferase (GGT), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) by automatic clinical chemistry analyzer (Cobus-Mira-Plus; Roche Diagnostic System, USA).
Detection of pro-inflammatory cytokine gene expression by real-time PCR
LPS challenge
Pigs were challenged with lipopolysaccharide (E coli 055:B5; Sigma, USA) 500 µg/kg BW by venae auriculares anteriores injection, and sacrificed after 3 h. The real-time PCR method was used to monitor the mRNA expressions of IL-2, IFN-γ, TNF-α, and β-actin.
Total RNA and real-time PCR
The total RNA from spleen tissue (0.2 mg) was extracted by the TRIZOL Reagent method (Invitrogen, USA) and reverse transcription was performed. The cytokine oligonucleotides listed in Table 2 were applied to amplify specific cDNA.
Real-time PCR was performed in a 7000 Fluorescence Quantitative PCR Cycler (Applied Biosystems, USA) starting with a 2 min UNG incubation step at 50℃ and 10 min AmpliTaq Gold Activation at 95℃, followed by a 2-step temperature cycling (15 sec at 95℃, 1 min at 60℃) and 35 cycles to complete polymerization.
Results
Blood biochemistry
Using blood biochemical parameters for estimation of toxic effects is based on the assumption that elevated activities of serum enzymes such as ALT and AST might reflect organ damage [4]. In this study, the total proteins, albumin, globulin, GGT, AST, and ALT were assayed in blood from both the control group and the toxin group. As shown in Table 3, the animals fed with DON and ZON showed lower total protein, albumin, and globulin than the control group, with decreases (p < 0.05) of 14.5%, 6.5% and 11.3%, respectively. Furthermore, DON and ZON induced higher enzyme activities of GGT, AST, and ALT with increases (p < 0.05) of 72.0%, 32.6% and 36.6%, respectively. All measured enzymatic activities of the control group were in the normal ranges of GGT (10-60 U/l), ALT (31-58 U/l), and AST (32-84 U/l) [7]. It is interesting to note that both albumin/globulin and AST/ALT ratios in the two groups showed no significant difference.
Immune responses
The titers of specific antibodies after vaccination are a good indication of humoral immunoresponses. As shown in Table 4, DON and ZON impaired the production of anti-CSF antibodies as the anti-CSF titers from the toxin group are significantly lower than those from the control group at the 28th day.
The real-time PCR results of mRNA expressions of IFN-γ, TNF-α, and IL-2 in the spleens of pigs are illustrated in mean ± SE of these results for different treatment (n = 12). Fig. 1. All data were expressed as relative mRNA expression of β-actin. As shown in Fig. 1, DON and ZON decreased the expression levels of all three cytokines tested; IFN-γ, TNF-α, and IL-2 had a decrease (p < 0.05) of 36.0%, 29.0%, and 35.4%, respectively.
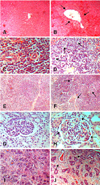
Histopathological examination
The histopathological changes caused by DON and ZON (Fig. 2). The pathological changes had multi-organ toxic characteristics including liver, spleen, lymph nodes, uterus, and kidney. No histopathological alterations were observed in the control group.
Discussion
In this study, we attempted to develop an experimental model for chronic mycotoxicosis in pigs as a result of the ingestion of a mixture of DON and ZON. From our year-long collection and analysis of feeds from pig farms, we concluded that there are approximately 1 mg/kg DON and 250 µg/kg ZON in diets used by pig farms in Southern China. According to the BML [2], this level of the two toxins is safe for starting and finishing pigs. But our data showed that supplementation of 1 mg/kg DON and 250 µg/kg ZON in diets had wide-ranging pathological effects in pigs.
It has been reported that elevated activities of serum enzymes such as ALT and AST might reflect organ damage [4]. A similar experiment demonstrated that total serum protein significantly decreased by 60% when animals were challenged by contaminated maize with Fusarium toxin (0.42 mg/kg ZON and 3.9 mg/kg DON) [5]. A prior paper reported an elevated serum albumin/globulin ratio due to increased serum albumin and decreased serum α-globulin concentrations in pigs fed with Fusarium mycotoxin-contaminated grains [15]. However, our study showed that the albumin/globulin ratio was not changed, even though the concentrations of both albumin and globulin decreased. It is possible that DON and other Fusarium mycotoxins directly affect globulin synthesis in the liver and compromise the immune response of pigs [15].
Altered GGT activities and urea concentrations were earlier observed in livestock and poultry fed contaminated grains, and might indicate that Fusarium mycotoxin induced hepatotoxicity. However, Kubena et al. [11] reported no effect on blood serum enzymes in chickens fed up to 16 mg/kg DON feed. This difference may be due to the different animal species used and the duration of the DON challenge. Our results showed that GGT, AST, and ALT activities increased after DON exposure for 6 weeks. In addition, the histopathological observations also confirmed the occurrence of liver and other organ damage.
Rotter et al. [15] reported a lower skin temperature, poorer feed efficiency, more corrugated stomach, reduced α-globulin levels and lower antibody titers to sheep red blood cells, in pigs consuming Fusarium mycotoxin-contaminated diet when compared with the pair-fed control pigs. Overnes et al. [12]observed a significant decrease in the secondary, but not in the primary, antibody responses to tetanus toxoid in pigs fed Fusarium mycotoxin-contaminated wheat. In our study the serum anti-CSF titer changes demonstrated that DON and ZON decreased the response of pigs to CSF vaccination. The antibody titers were significantly decreased at the 28th day in the toxin group compared to those of the control, although there was no significant difference in antibody titers at the 14th and 42nd day.
It has been reported that a single oral exposure to both 5 and 25 mg DON/kg BW of mice significantly induces the mRNAs for the proinflammatory cytokines interleukin (IL)-1β, IL-6, and tumor necrosis factor-α; the T helper 1 cytokines interferon-γ and IL-2; and the T helper 2 cytokines IL-4 and IL-10, whereas lower doses had no effect [19]. Any of these cytokines could directly or indirectly enhance differentiation of IgA-secreting B cells. However, when mice were fed sub-chronic levels of DON (0, 10, and 25 mg/kg) for 4 weeks, increased mRNA expression was most prominent for IL-2, interferon-γ, IL-10, and tumor necrosis factor-α [20]. Similar results also demonstrate that DON (25 mg/kg BW) induces gene expression of IL-1α, IL-1β, IL-6, and IL-11 in mice [10]. In most of these published papers animals were challenged by a dosage of DON which was much higher than the toxin level used in the present experiment. As in the present study domestic animals were exposed to mixtures of fungal toxins under field conditions. Therefore, the present results could reflect the practical situation in the field with mild toxin levels and longer-term (6 weeks) exposure, and show that the gene expression of selected cytokines was impaired. However, the combined effects of DON and ZON on the mechanisms of toxicity of the potential immunotoxins are still unknown.
The histopathological changes in the DON and ZON challenged group showed the characteristics of multiorgan toxicity including liver, spleen, lymph node, uterus, and kidney pathology. Trichothecene mycotoxins, like DON, binds to ribosomal peptidyl-transferase and specifically inhibits protein and DNA synthesis, so exposure results in decreased cell proliferation [16]. The cytotoxicity of DON in the liver has not been reported in the field and cases of DON intoxication in domestic animals have not been experimental. This may be due to the remarkable potential of the liver for regeneration and rapid clearance of apoptotic cells in vivo. Ihara et al. [8] reported that apoptosis was induced more rapidly by T-2 toxin in the liver than in other tissues observed in vivo, and was detectable in the liver at 2 h but not 12 h later. DON is less toxic than T-2 toxin, but the same process may occur in DON intoxication. In the present experiment, only blood vessel thickening and dilatation were found in the liver sections. The effect of DON and ZON on the uterus including congestion or hyperemia, and blood vessel dilatation was seen in our experiment. The previous report indicates that 1 mg/kg is the minimum concentration to produce hyperestrogenism [9]. From our research it would seem that ZON levels of 250 µg/kg will produce a hyper-estrogenic appearance in female pigs. Decreased concentrations of serum protein and albumin were observed in the present study. Bergsjo et al. [1] also consider that these effects may be secondary to the reduced feed uptake, but inhibition of protein synthesis by DON in the liver may play some role. One of the toxicities of DON is thought to derive from the inhibition of protein synthesis [13].
Overall, our results showed that supplementation of 1 mg/kg DON and 250 µg/kg ZON in diets caused wide ranging pathological effects in pigs. It should be noted that the concentrations of DON and ZON used in this study are in line with the published critical values permitted by BML [2]. Thus our study clearly puts the standard and adequacy of the safety measures for these toxins into question. The authors consider that with the increasing availability of cellular and molecular technologies, it is time to revisit the safety standards for toxins in feeds so that we make our feeds safer, providing our consumers with safer products.





 XML Download
XML Download